
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 322
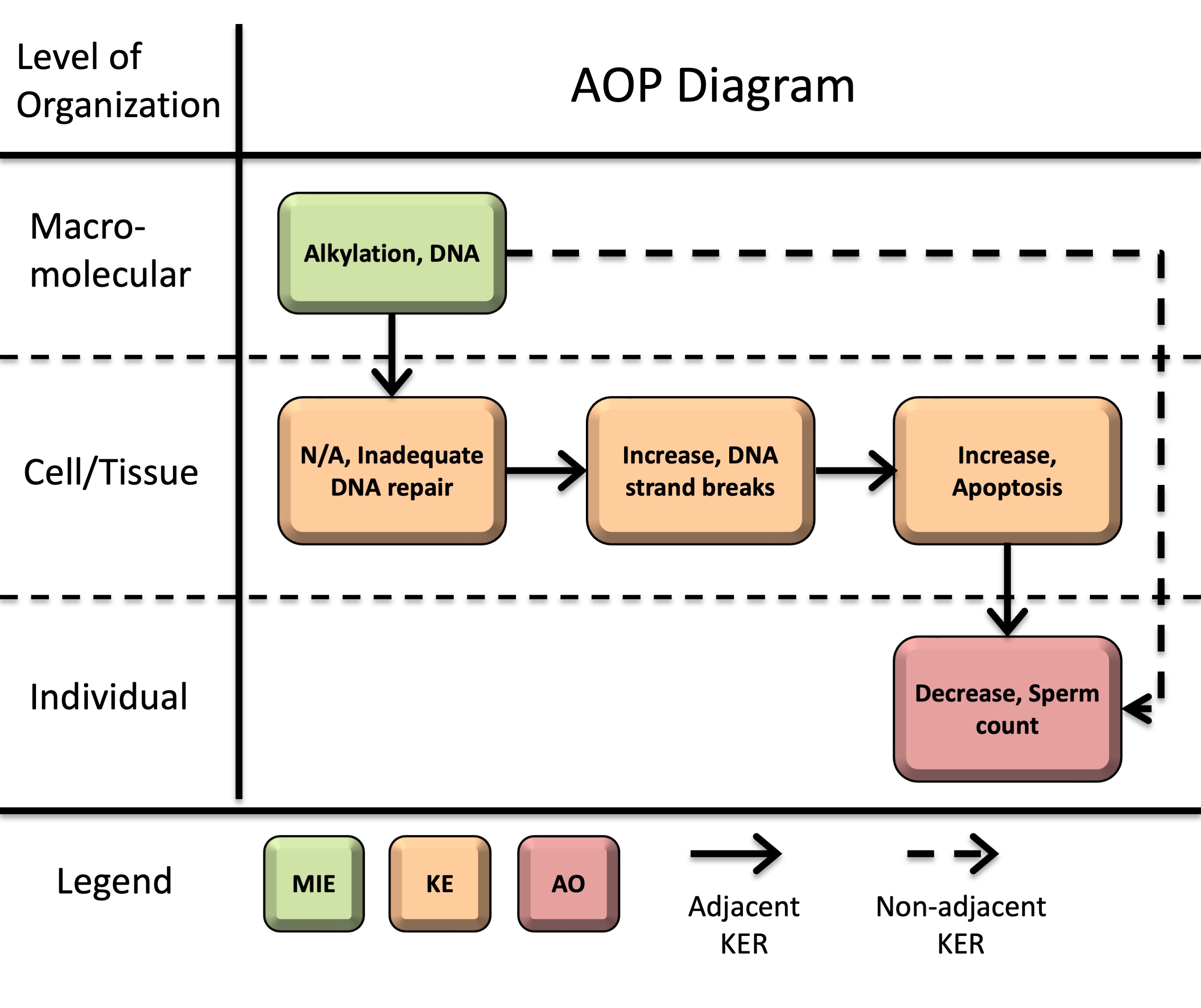
Title
Alkylation of DNA leading to decreased sperm count
Short name
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 322, Alkylation of DNA leading to decreased sperm count in tools offered by third parties.
Point of Contact
Contributors
- Carole Yauk
- Xiaotong Wang
- Francesco Marchetti
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on July 05, 2026 15:29
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Alkylation, DNA | September 16, 2017 10:14 |
| Inadequate DNA repair | March 08, 2024 12:15 |
| Increase, DNA strand breaks | December 17, 2024 11:57 |
| Decrease, Sperm count | July 02, 2026 23:33 |
| Apoptosis | May 31, 2025 08:50 |
| Alkylation, DNA leads to Decrease, Sperm count | July 02, 2026 23:43 |
| Alkylation, DNA leads to Inadequate DNA repair | December 10, 2019 10:43 |
| Inadequate DNA repair leads to Increase, DNA strand breaks | July 03, 2026 13:37 |
| Increase, DNA strand breaks leads to Apoptosis | July 03, 2026 13:08 |
| Apoptosis leads to Decrease, Sperm count | July 03, 2026 13:12 |
Abstract
Decreased sperm count is a key endpoint in the assessment of male reproductive health as it is directly associated with impaired fertility. Exposure to DNA alkylating agents, including chemotherapeutic drugs and environmental toxicants, is associated with reduced sperm counts in experimental models and humans. However, the progression from DNA lesions to reduced sperm output has not been systematically organized in an AOP framework. This AOP addresses that gap by describing how DNA alkylation can lead to inadequate DNA repair, increased DNA strand breaks, apoptosis, impaired spermatogenesis, and ultimately decreased sperm count. Although genotoxicity data are not routinely used as predictors of male fertility effects, this AOP provides a basis for evaluating when such data may be informative for reproductive toxicity assessment and for developing future predictive toxicology approaches.
This AOP initiates with DNA alkylation (molecular initiating event, MIE). Alkylation-induced DNA lesions can then overwhelm DNA repair capacity (key event, KE1: inadequate repair) and an accumulation of DNA strand breaks (KE2). Persistent or unrepaired DNA damage activates DNA damage response pathways, ultimately leading to apoptosis (KE3). When apoptosis occurs in male germ cells and supporting testicular cells, excessive depletion of the developing germ cell population and disruption of structural support and endocrine signaling in the testis lead to decreased sperm counts (adverse outcome, AO) in sexually mature males.
This pathway is supported by strong biological plausibility and moderate to strong empirical evidence across multiple model systems, including human data, though quantitative understanding remains limited.
AOP Development Strategy
Context
The development of this AOP was motivated by the need to organize the well-established relationship between DNA alkylation and impaired male reproductive function into a formal mechanistic framework. DNA alkylation is a well-characterized form of genotoxic damage (Soll et al., 2017). In male spermatogonia and meiotic cells, alkylation of DNA in actively proliferating germ cells can trigger DNA damage responses, cell cycle arrest, and apoptosis, ultimately impairing spermatogenesis and leading to adverse reproductive outcomes (Kaina, 2003; Rübe et al., 2011; Li et al., 2025). Exposure to alkylating agents, particularly in the context of cancer chemotherapy, has long been associated with reduced sperm counts, oligozoospermia, azoospermia, and impaired fertility in males, with severity and recovery largely dependent on cumulative dose (reviewed by Howell and Shalet, 2005; Okada and Fujisawa, 2018). However, genotoxicity data are not routinely used as predictors of male fertility effects in reproductive toxicity assessment. This creates a need for a structured framework to evaluate when DNA damage in the male germline may be informative for reproductive hazard.
Human studies have reported associations between biomarkers of DNA alkylation and reduced sperm concentrations (Altakroni et al., 2021). Evidence from childhood cancer survivors also demonstrates that exposure during early life can impair germ cell populations and lead to reduced sperm production later in adulthood (Beaud et al., 2019; reviewed by Delessard et al., 2019). Experimental studies further demonstrate that alkylating agents produce dose-dependent and persistent reductions in sperm counts across species, including rodents, non-human primates, and humans (Meistrich, 1982a; Bucci and Meistrich, 1987; Hermann et al., 2009). Although DNA alkylation may occur during fetal, juvenile, or adult life stages, and can impair germ cell populations at any of these stages, the downstream events in this AOP involve spermatogenesis and sperm production; thus, the adverse outcome is manifested in sexually mature males.
This AOP branches from an existing AOP developed by Yauk et al., “Alkylation of DNA in Male Premeiotic Germ Cells Leading to Heritable Mutations” (AOP15), and contributes to the development of a broader AOP network for genotoxicity and reproductive toxicity.
An additional objective of this work is to facilitate the use of new approach methods (NAMs) in regulatory decision-making. By mechanistically linking early biological responses and adverse outcomes of regulatory concern, this AOP supports the development of novel models and screening tools to identify chemicals that may impair male fertility and provides a context for the use of data from NAMs. Additionally, by systematically organizing the existing knowledge on this topic we have identified key data gaps to guide future research in the field.
Strategy
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 97 | Alkylation, DNA | Alkylation, DNA |
| KE | 155 | Inadequate DNA repair | Inadequate DNA repair |
| KE | 1635 | Increase, DNA strand breaks | Increase, DNA strand breaks |
| KE | 1262 | Apoptosis | Apoptosis |
| AO | 1757 | Decrease, Sperm count | Decrease, Sperm count |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Alkylation, DNA leads to Inadequate DNA repair | adjacent | High | Moderate |
| Inadequate DNA repair leads to Increase, DNA strand breaks | adjacent | High | Moderate |
| Increase, DNA strand breaks leads to Apoptosis | adjacent | High | Moderate |
| Apoptosis leads to Decrease, Sperm count | adjacent | High | Low |
| Alkylation, DNA leads to Decrease, Sperm count | non-adjacent | High | Low |
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Juvenile | High |
| Prepubertal | High |
| Adult, reproductively mature | High |
| Fetal | Moderate |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Overall Assessment of the AOP
Domain of Applicability
DNA alkylation can occur in all cell types. DNA repair processes and apoptotic pathways are conserved across species. While decreased sperm counts are measurable only after sexual maturity, the upstream KEs and KERs are biologically plausible and operative across fetal, juvenile, and adult life stages. Therefore, the overall biological domain of applicability of the AOP is considered relevant to male individuals exposed during fetal, juvenile, or adult stages, with manifestation of the AO occurring after reproductive maturation. In the male reproductive system, this AOP is most relevant when alkylation damage occurs in proliferating or meiotic germ cells that contribute directly to sperm production.
Essentiality of the Key Events
Overall, the essentiality of KEs in this AOP is supported primarily by indirect evidence from studies involving genetic manipulation, pharmacological intervention, and endpoint recovery, demonstrating that perturbation of upstream KEs is associated with corresponding changes in downstream KEs. A summary of the evidence supporting the essentiality of individual KEs and corresponding uncertainties or inconsistencies are provided in Table 1: summary of supporting evidence for essentiality of key events.
Impairment of DNA repair and DNA damage response (DDR) pathways consistently results in increased DNA strand breaks (KE2) and downstream apoptosis (KE3) in the presence of alkylating agents, indicating that upstream KEs are required for progression of the pathway. Although simultaneous measurements of alkyl DNA adducts, repair capacity, and sperm production within the same study are limited, exposure studies involving alkylating agents consistently show that greater DNA damage is associated with more severe and persistent impairment of sperm production across species, including rodents, non-human primates, and humans (Meistrich, 1982a, 1982b; Bucci and Meistrich, 1987; Hermann et al., 2009, Meistrich et al., 1992; Howell and Shalet, 2005; Okada and Fujisawa, 2018; Beaud et al., 2019). Together, these findings support the functional importance of DNA alkylation and DDR-related processes in progression of this AOP.
Essentiality of DNA strand breaks (KE2) is supported by studies showing that disruption of DDR signaling pathways (e.g., ATM inhibition) can prevent apoptosis even when DNA damage is present (Rodrigues et al., 2013). These findings highlight that damage sensing and downstream DDR signaling are required for the pathway to progress toward apoptotic cell death.
Evidence supporting the essentiality of apoptosis (KE3) is provided by intervention studies showing that attenuation of apoptotic signaling is associated with recovery of sperm counts (AO). Concordant reversibility across upstream and downstream endpoints following chemical or biological intervention supports the contribution of apoptosis to the AO (Oyovwi et al., 2023; Yaman et al., 2018; Udefa et al., 2020; Ehghaghi et al., 2022). However, many protective interventions also modulate oxidative stress and inflammatory pathways simultaneously, making it difficult to isolate apoptosis as the sole driver of sperm recovery.
Uncertainties include limitations in study design (e.g., reliance on single collection timepoints that do not capture temporal progression or delayed changes in sperm output), assay specificity (e.g., distinguishing primary DNA strand breaks from apoptotic DNA fragmentation), and incomplete characterization of quantitative relationships. Nonetheless, the overall weight of evidence supports a moderate level of essentiality.
Table 1. Summary of Supporting Evidence for Essentiality of Key Events
|
Event |
Direct Essentiality Evidence |
Indirect Essentiality Evidence |
Uncertainties or Inconsistency |
|
DNA alkylation (MIE) |
Limited direct evidence |
Exposure to alkylating agents leads to dose-dependent loss of germ cells and subsequent reductions in sperm counts (AO) across species; recovery may occur following removal of the stressor In rodents, treatment with alkylating agents at increasing doses results in dose-dependent decreases in testicular or epididymal sperm counts (Meistrich, 1982a, 1982b; Bucci and Meistrich, 1987). In rhesus macaques, similar progressive, dose-dependent declines have been observed following busulfan exposure, with higher doses leading to more persistent reductions in sperm production (Hermann et al., 2009). In cancer patients, the use of DNA alkylating drugs is strongly associated with lower sperm counts; such link is not observed in patients receiving non-alkylating drugs (Beaud et al., 2019). Repeated exposure to an alkylating agent caused a marked reduction in sperm counts in mice, with gradual recovery following cessation of exposure, demonstrating reversibility of the AO after removal of the stressor (Yin et al., 2014). |
Formation of alkyl DNA adducts in germ cells has been demonstrated in vivo; however, there are limited integrated measurements of the MIE, downstream KEs, the AO in the same studies. Consequently, progression through the pathway is often inferred from the known mechanisms of alkylating agents. |
|
Inadequate repair (KE1) |
Depletion of O6-alkylguanine-DNA alkyltransferase (AGT/MGMT) leads to corresponding alterations in DNA strand break (KE2) formation Key studies: Roos et al. (2004) linked DNA alkylation (MIE), impaired repair (KE1), DNA strand breaks (KE2), and apoptosis (KE3) in proliferating human lymphocytes. Inactivation of MGMT increases persistence of alkylation-induced lesions, resulting in replication-dependent strand break formation and subsequent apoptosis. Carlsson et al. (2025) showed that pharmacological inhibition of MGMT enhanced N-nitrosodimethylamine-induced formation of DNA adducts (MIE), DNA strand breaks (KE2), and micronucleus formation in HepG2-CYP2E1 human liver cells. |
Modulation of DNA repair (KE1) or DDR pathways leads to concurrent increases in DNA strand breaks (KE2) and apoptosis (KE3) Knockdown or knockout of CDKN2AIP, a regulator of DNA repair, leads to increased double strand breaks (DSBs) and apoptosis in mouse Sertoli cells and male germ cells (Cao et al., 2022). Greater impairment of DDR pathways (Nbn/Atm double deletion) results in more DSBs and apoptotic cells than Nbn single deletion in mouse neuronal tissues (Rodrigues et al., 2013). Similarly, single or double deletion of Apc/p53 (Méniel et al., 2015), bidirectional genetic modulation of CIRKIL/Ku70 (Xiao et al., 2023), inhibition of the PI3K/mTOR pathway (Liu et al., 2014), or homologous recombination (Stringer et al., 2020), result in more DSBs and apoptosis than wildtype or single deletion models. |
DNA repair capacity is not measured directly in many studies. Accumulation of DSBs following impairment of DDR pathways is interpreted as evidence of insufficient repair. |
|
DNA strand breaks (KE2) |
Blocking signaling downstream of DNA strand breaks (KE2) prevents apoptosis (KE3) ATM inactivation prevents apoptosis in eye retina, despite the presence of DSBs (Rodrigues et al., 2013). |
Modulation of the magnitude of DNA strand breaks (KE2) is associated with a corresponding change in apoptosis (KE3) The intervention studies listed in indirect evidence for KE1 also demonstrate a graded response-response relationship between DSBs and apoptosis. |
TUNEL staining may detect both primary strand breaks and apoptotic DNA fragmentation, when KE2 and KE3 are measured at overlapping timepoints. More specific DSB markers (e.g., γH2AX) were used in several studies. The absence of detectable DNA strand breaks may reflect limitations in the sampling time and assay specificity/sensitivity (e.g., the alkaline comet assay may miss DSBs, or the 24-hour sampling window may miss transient, repaired lesions). |
|
Apoptosis (KE3) |
Limited direct evidence |
Attenuation of apoptosis (KE3) leads to recovery of sperm counts (AO) Key study (Oyovwi et al., 2023): Pharmacological attenuation of apoptosis using quercetin fully reversed testicular damage and restored sperm counts following levetiracetam exposure in rats. Multiple KEs were measured in this study, including sperm DNA fragmentation index (KE2; inferred from aniline blue staining), apoptotic markers (KE3: caspase-3, p53, cytochrome c, Bcl-2), and the AO (testicular sperm counts and histological evidence of germ cell loss). The concordant reversal of these endpoints following the intervention provides strong support for the progression across KEs. Additional intervention studies using antioxidants or protective agents (e.g., L-carnitine, plant extracts, quercetin, selenium nanoparticles, and probiotics) demonstrate that reducing apoptotic signaling is associated with improved sperm counts following exposure to chemotherapeutic agents, toxicants, or radiation (Yaman et al., 2018; Udefa et al., 2020; Ehghaghi et al., 2022). |
Protective agents often suppress inflammation and oxidative stress simultaneously. It is unclear if sperm recovery is due to reduced apoptosis or these co-activated pathways, or both. Apoptotic markers are often measured in whole testis homogenates and the AO is likely caused by apoptosis of mixed testicular cell populations. Inappropriate sampling time and high variability in sperm count data may lead to "false negatives" (Gur et al., 2023). |
Evidence Assessment
|
Biological plausibility of KERs |
Defining question |
High (Strong) |
Moderate |
Low (Weak) |
|
Is there a mechanistic relationship between KEup and KEdown consistent with established biological knowledge? |
Extensive understanding of the KER based on extensive previous documentation and broad acceptance. |
KER is plausible based on analogy to accepted biological relationships, but scientific understanding is incomplete. |
Empirical support for association between KEs, but the structural or functional relationship between them is not understood. |
|
|
MIE KE1: Alkylation, DNA leads to inadequate DNA repair |
STRONG Extensive evidence indicates that sufficiently high level of DNA alkylation can overwhelm cellular DNA repair machinery, leading to the persistence of DNA adducts and other unrepaired lesions. AGT, also known as MGMT in mammals, is an established suicide enzyme that can become saturated at high doses or after repeated exposure to alkylating agents, leading to inadequate DNA repair and accumulation of DNA alkyl adducts. This relationship is broadly conserved across species and cell types. |
|||
|
KE1 KE2: Inadequate DNA repair leads to Increase, DNA strand breaks |
STRONG DNA adducts and repair intermediates can accumulate when alkylation damage exceeds repair capacity in cells, including the depletion of AGT/MGMT. It is well established that persistence of unrepaired alkyl DNA lesions can interfere with DNA replication and promote replication fork stalling, leading to DNA strand breaks, which then activate DDR pathways. Extensive mechanistic evidence supports a causal relationship between inadequate DNA repair of alkylation-induced damage and increased DNA strand breaks. |
|||
|
KE2 KE3: Increase, DNA strand breaks leads to Apoptosis |
STRONG There is extensive mechanistic understanding of the DNA damage response pathways that link DNA strand breaks and apoptosis through both p53-dependent and independent mechanisms. |
|||
|
KE3 AO: Apoptosis leads to Decrease, Sperm Count |
STRONG Loss of testicular cells (e.g., developing germ cells and supportive somatic cells) through apoptosis disrupts normal testicular function to support spermatogenesis, resulting in a subsequent decrease in mature sperm output. These mechanisms are well established across mammalian systems. |
|||
|
MIE AO: Alkylation, DNA leads to Decrease, Sperm Count |
STRONG The mechanistic linkage is conserved across species and supported by extensive knowledge of germ cell biology and toxicology. While alkylation damage can occur across all stages of spermatogenesis, effects on sperm counts are primarily driven by damage to proliferating and meiotic germ cells, whereas damage to post-meiotic cells predominantly affects sperm quality rather than quantity. |
|||
|
Essentiality of KEs |
Defining question |
High (Strong) |
Moderate |
Low (Weak) |
|
Are downstream KEs and/or the AO prevented if an upstream KE is blocked? |
Direct evidence from specifically designed experimental studies illustrating essentiality for at least one of the important KEs. |
Indirect evidence that sufficient modification of an expected modulating factor attenuates or augments a KE. |
No or contradictory experimental evidence of the essentiality of any of the KEs. |
|
|
AOP-level |
MODERATE Evidence supporting the essentiality of KEs is available from genetic and mechanistic studies. Modulation of DNA damage response and apoptotic pathways induce corresponding changes in downstream outcomes, including apoptosis and sperm counts. A limited number of studies provide more direct evidence of essentiality for specific KERs, while identifying a few essential signaling mediators involved in the transduction of DNA damage into apoptosis. However, such direct evidence is not consistently available across all KEs in the pathway, and much of the support remains indirect or context-specific. |
|||
|
Empirical support for KERs |
Defining question |
High (Strong) |
Moderate |
Low (Weak) |
|
Does empirical evidence support that a change in KEup leads to an appropriate change in KEdown? Does KEup occur at lower doses and earlier time points than KE down and is the incidence of KEup> than that for KEdown? Inconsistencies? |
Multiple studies showing dependent change in both events following exposure to a wide range of specific stressors. No or few critical data gaps or conflicting data. |
Demonstrated dependent change in both events following exposure to a small number of stressors. Some inconsistencies with expected pattern that can be explained by various factors. |
Limited or no studies reporting dependent change in both events following exposure to a specific stressor; and/or significant inconsistencies in empirical support across taxa and species that don’t align with hypothesized AOP. |
|
|
MIE KE1: Alkylation, DNA leads to inadequate DNA repair |
STRONG Inadequate DNA repair is measured indirectly through persistence of DNA adducts or increases in mutations that result from unrepaired DNA damage. Extensive evidence from somatic and germ cells supports this KER (in particular for temporal concordance), although quantitative dose-response concordance is less well characterized. There are no apparent inconsistencies. Numerous studies demonstrate that alkyl DNA adducts persist when repair capacity is exceeded or repair pathways are impaired. In particular, saturation or depletion of AGT/MGMT results in increased persistence of O6-alkylguanine adducts, providing strong support for this KER. |
|||
|
KE1 KE2: Inadequate DNA repair leads to Increase, DNA strand breaks |
MODERATE Limited in vivo data are available. However, multiple in vitro and genetic studies demonstrate that impairment of DNA repair or DDR pathways result in increased accumulation and persistence of DNA strand breaks in both somatic cells and germ cells following exposure to genotoxic stressors. In the context of DNA alkylation, saturation or depletion of AGT/MGMT leads to persistence of O6-alkylguanine lesions, which are subsequently converted into DNA strand breaks, consistent with temporal concordance. Studies involving AGT depletion and repair-deficient systems provide empirical support for the essential role of inadequate repair of alkylation DNA damage in the accumulation of DNA strand breaks. |
|||
|
KE2 KE3: Increase, DNA strand breaks leads to Apoptosis |
STRONG Temporal concordance is consistently observed, with DNA strand breaks occurring earlier or concurrently with apoptotic responses across in vitro somatic and germ cells, and rodent models. Dose concordance is supported, although dose-response data are limited in some studies. Evidence for incidence concordance is supported by a small number of studies, while others are limited by lack of appropriate measurements. |
|||
|
KE3 AO: Apoptosis leads to Reduce, Sperm Count |
STRONG Concordant changes between increased apoptosis and decreased sperm counts are have been consistently observed across multiple in vivo rodent studies. Temporal alignment is biologically supported, although it is often inferred rather than directly measured. Evidence for dose concordance is limited as many studies used a single exposure dose, preventing assessment of dose-dependent changes. Incidence concordance is generally not assessed, as both apoptosis and sperm count are typically reported as continuous outcomes (e.g., group means) rather than as the proportion of individual animals meeting predefined criteria for increased apoptosis or reduced sperm counts. Nevertheless, the consistency of the empirical evidence, together with multiple intervention studies demonstrating recovery of sperm counts following attenuation of apoptotic signaling, provides strong support for this KER. |
|||
|
MIE AO: Alkylation, DNA leads to Decrease, Sperm Count |
STRONG Empirical evidence from both experimental animal models and human studies supports a consistent relationship between DNA alkylation and reduced sperm counts. Although direct measurement of both KEs in the same study is limited, extrapolation across studies involving exposure to well-characterized alkylating agents provides strong empirical support for temporal and dose concordance. Reduction in sperm counts occur after delays consistent with spermatogenic progression, and higher exposures lead to greater and more sustained decreases in sperm counts across multiple studies and stressors. |
|||
Known Modulating Factors
|
Modulating Factor |
Influence on Outcome |
KER(s) involved |
|
DNA repair capacity |
Determines the threshold at which DNA alkylation damage exceeds repair capacity and leads to insufficient or incorrect repair; lower capacity increases persistence and numbers of DNA lesions and the severity of downstream effects. |
MIEKE1 KE1KE2 KE2KE3 |
|
Exposure dose/duration |
Exposure doses and duration are strong modulating factors of the magnitude of effects across KEs. Higher doses or prolonged exposure are expected to increase cumulative DNA damage, which can lead to more severe downstream effects. At overtly cytotoxic concentrations (i.e., >50%, in the context of in vitro genotoxicity testing), non-specific cytotoxicity can occur, making it difficult to attribute apoptotic responses and the AO specifically to DNA alkylation. |
All KERs |
|
Stress-related pathways |
Co-activation of parallel stress response pathways, oxidative stress, and/or inflammation, can amplify DNA damage and apoptosis, enhancing downstream effects independent of DNA alkylation. |
KE1KE2 KE2KE3 |
|
Germ cell developmental stage |
Proliferating or differentiating germ cells are more susceptible to DNA alkylation damage. Damage in spermatogonial stem cells (SSCs) leads to long-term or permanent reductions in sperm counts; damage in later stages results in transient effects or reduces sperm quality rather than quantity. |
KE2KE3 KE3AO MIEAO |
|
Spermatogenic kinetics |
The delay between DNA damage and observable reduction in sperm counts, as well as the time required for recovery (if SSCs survive), is determined by species-specific spermatogenic kinetics. |
KE3AO MIEAO |
Quantitative Understanding
The overall quantitative understanding of the KERs in this AOP is low. While individual KERs are supported by qualitative evidence of dose-response and temporal concordance, quantitative relationships between KEs are not well defined.
Some studies demonstrate graded changes between adjacent KEs following genetic or pharmacological modulation (e.g., KER2 and KER3), supporting response-response relationships. More details are provided in the individual KERs.
A threshold-based response is expected in this AOP, as DNA damage must exceed the repair capacity to propagate to downstream effects. In addition, a sufficient level of germ cell apoptosis is likely required before a measurable decline in sperm count occurs. However, several modulating factors have been identified, and the quantitative relationships are not generalizable across cell types, tissues, or developmental stages.
Considerations for Potential Applications of the AOP (optional)
References
Altakroni, B., Nevin, C., Carroll, M., Murgatroyd, C., Horne, G., Brison, D. R. & Povey, A. C. (2021). The marker of alkyl DNA base damage, N7-methylguanine, is associated with semen quality in men. Scientific Reports, 11(1), 3121. https://doi.org/10.1038/s41598-021-81674-x
Beaud, H., Albert, O., Robaire, B., Rousseau, M. C., Chan, P. T. K. & Delbes, G. (2019). Sperm DNA integrity in adult survivors of paediatric leukemia and lymphoma: A pilot study on the impact of age and type of treatment. PLoS ONE, 14(12), e0226262. https://doi.org/10.1371/journal.pone.0226262
Bucci, L. R. & Meistrich, M. L. (1987). Effects of busulfan on murine spermatogenesis: cytotoxicity, sterility, sperm abnormalities, and dominant lethal mutations. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, 176(2), 259–268. https://doi.org/10.1016/0027-5107(87)90057-1
Carlsson, M. J., Herzog, N., Felske, C., Ackermann, G., Regier, A., Wittmann, S., Cereijo, R. F., Sturla, S. J., Küpper, J.-H. & Fahrer, J. (2025). The DNA Repair Protein MGMT Protects against the Genotoxicity of N‑Nitrosodimethylamine, but Not N‑Nitrosodiethanolamine and N‑Nitrosomethylaniline, in Human HepG2 Liver Cells with CYP2E1 Expression. Chemical Research in Toxicology, 38(6), 1134–1146. https://doi.org/10.1021/acs.chemrestox.5c00133
Cao, Y., Sun, Q., Chen, Z., Lu, J., Geng, T., Ma, L. & Zhang, Y. (2022). CDKN2AIP is critical for spermiogenesis and germ cell development. Cell & Bioscience, 12(1), 136. https://doi.org/10.1186/s13578-022-00861-z
Delessard, M., Saulnier, J., Rives, A., Dumont, L., Rondanino, C. & Rives, N. (2020). Exposure to Chemotherapy During Childhood or Adulthood and Consequences on Spermatogenesis and Male Fertility. International Journal of Molecular Sciences, 21(4), 1454. https://doi.org/10.3390/ijms21041454
Ehghaghi, A., Zokaei, E., Modarressi, M. H., Tavoosidana, G., Ghafouri-Fard, S., Khanali, F., Motevaseli, E. & Noroozi, Z. (2022). Antioxidant and anti-apoptotic effects of selenium nanoparticles and Lactobacillus casei on mice testis after X-ray. Andrologia, 54(11), e14591. https://doi.org/10.1111/and.14591
Gur, C., Akarsu, S. A., Akaras, N., Tuncer, S. C. & Kandemir, F. M. (2023). Carvacrol reduces abnormal and dead sperm counts by attenuating sodium arsenite-induced oxidative stress, inflammation, apoptosis, and autophagy in the testicular tissues of rats. Environmental Toxicology, 38(6), 1265–1276. https://doi.org/10.1002/tox.23762
Hermann, B. P., Sukhwani, M., Lin, C., Sheng, Y., Tomko, J., Rodriguez, M., Shuttleworth, J. J., McFarland, D., Hobbs, R. M., Pandolfi, P. P., Schatten, G. P. & Orwig, K. E. (2009). Characterization, Cryopreservation, and Ablation of Spermatogonial Stem Cells in Adult Rhesus Macaques. Stem Cells, 25(9), 2330–2338. https://doi.org/10.1634/stemcells.2007-0143
Howell, S. J. & Shalet, S. M. (2005). Spermatogenesis After Cancer Treatment: Damage and Recovery. JNCI Monographs, 2005(34), 12–17. https://doi.org/10.1093/jncimonographs/lgi003
Kaina, B. (2003). DNA damage-triggered apoptosis: critical role of DNA repair, double-strand breaks, cell proliferation and signaling. Biochemical Pharmacology, 66(8), 1547–1554. https://doi.org/10.1016/s0006-2952(03)00510-0
Li, N., Wang, H., zou, S., Yu, X. & Li, J. (2025). Perspective in the Mechanisms for Repairing Sperm DNA Damage. Reproductive Sciences, 32(1), 41–51. https://doi.org/10.1007/s43032-024-01714-5
Liu, W.-L., Gao, M., Tzen, K.-Y., Tsai, C.-L., Hsu, F.-M., Cheng, A.-L. & Cheng, J. C.-H. (2014). Targeting Phosphatidylinositide3-Kinase/Akt pathway by BKM120 for radiosensitization in hepatocellular carcinoma. Oncotarget, 5(11), 3662–3672. https://doi.org/10.18632/oncotarget.1978
Meistrich, M. L. (1982a). Quantitative Correlation Between Testicular Stem Cell Survival, Sperm Production, and Fertility in the Mouse After Treatment With Different Cytotoxic Agents. Journal of Andrology, 3(1), 58–68. https://doi.org/10.1002/j.1939-4640.1982.tb00646.x
Meistrich, M. L., Finch, M., Cunha, M. F. da, Hacker, U. & Au, W. W. (1982b). Damaging effects of fourteen chemotherapeutic drugs on mouse testis cells. Cancer Research, 42(1), 122–131.
Meistrich, M. L., Wilson, G., Brown, B. W., Cunha, M. F. da & Lipshultz, L. I. (1992). Impact of cyclophosphamide on long-term reduction in sperm count in men treated with combination chemotherapy for Ewing and soft tissue sarcomas. Cancer, 70(11), 2703–2712. https://doi.org/10.1002/1097-0142(19921201)70:11<2703::aid-cncr2820701123>3.0.co;2-x
Méniel, V., Megges, M., Young, M. A., Cole, A., Sansom, O. J., Clarke, A. R. (2015). Apc and p53 interaction in DNA damage and genomic instability in hepatocytes. Oncogene, 34(31), 4118–4129. https://doi.org/10.1038/onc.2014.342
Murphy, C. J. & Richburg, J. H. (2015). Implications of Sertoli cell induced germ cell apoptosis to testicular pathology. Spermatogenesis, 4(2), e979110. https://doi.org/10.4161/21565562.2014.979110
Okada, K. & Fujisawa, M. (2018). Recovery of Spermatogenesis Following Cancer Treatment with Cytotoxic Chemotherapy and Radiotherapy. The World Journal of Men’s Health, 36(2), 166–174. https://doi.org/10.5534/wjmh.180043
Oyovwi, M. O., Oghenetega, O. B., Victor, E., Faith, F. Y. & Uchechukwu, J. G. (2023). Quercetin protects against levetiracetam induced gonadotoxicity in rats. Toxicology, 491, 153518. https://doi.org/10.1016/j.tox.2023.153518
Roos, W., Baumgartner, M. & Kaina, B. (2004). Apoptosis triggered by DNA damage O6-methylguanine in human lymphocytes requires DNA replication and is mediated by p53 and Fas/CD95/Apo-1. Oncogene, 23(2), 359–367. https://doi.org/10.1038/sj.onc.1207080
Rodrigues, P. M. G., Grigaravicius, P., Remus, M., Cavalheiro, G. R., Gomes, A. L., Rocha-Martins, M., Martins, M. R., Frappart, L., Reuss, D., McKinnon, P. J., Deimling, A. von, Martins, R. A. P. & Frappart, P.-O. (2013). Nbn and Atm Cooperate in a Tissue and Developmental Stage-Specific Manner to Prevent Double Strand Breaks and Apoptosis in Developing Brain and Eye. PLoS ONE, 8(7), e69209. https://doi.org/10.1371/journal.pone.0069209
Rübe, C. E., Zhang, S., Miebach, N., Fricke, A. & Rübe, C. (2011). Protecting the heritable genome: DNA damage response mechanisms in spermatogonial stem cells. DNA Repair, 10(2), 159–168. https://doi.org/10.1016/j.dnarep.2010.10.007
Soll, J. M., Sobol, R. W. & Mosammaparast, N. (2017). Regulation of DNA Alkylation Damage Repair: Lessons and Therapeutic Opportunities. Trends in Biochemical Sciences, 42(3), 206–218. https://doi.org/10.1016/j.tibs.2016.10.001
Udefa, A. L., Amama, E. A., Archibong, E. A., Nwangwa, J. N., Adama, S., Inyang, V. U., Inyaka, G. U., Aju, G. J., Okpa, S. & Inah, I. O. (2020). Antioxidant, anti-inflammatory and anti-apoptotic effects of hydro-ethanolic extract of Cyperus esculentus L. (tigernut) on lead acetate-induced testicular dysfunction in Wistar rats. Biomedicine & Pharmacotherapy, 129, 110491. https://doi.org/10.1016/j.biopha.2020.110491
Xiao, H., Zhang, M., Wu, H., Wu, J., Hu, X., Pei, X., Li, D., Zhao, L., Hua, Q., Meng, B., Zhang, X., Peng, L., Cheng, X., Li, Z., Yang, W., Zhang, Q., Zhang, Y., Lu, Y. & Pan, Z. (2022). CIRKIL Exacerbates Cardiac Ischemia/Reperfusion Injury by Interacting With Ku70. Circulation Research, 130(5), e3–e17. https://doi.org/10.1161/circresaha.121.318992
Yaman, O., & Topcu-Tarladacalisir, Y. (2018). L-carnitine counteracts prepubertal exposure to cisplatin induced impaired sperm in adult rats by preventing germ cell apoptosis. Biotechnic & Histochemistry, 1-11. doi:10.1080/10520295.2017.1401661
Yauk, C. L., Lambert, I. B., Meek, M. E. B., Douglas, G. R. & Marchetti, F. (2015). Development of the adverse outcome pathway “alkylation of DNA in male premeiotic germ cells leading to heritable mutations” using the OECD’s users’ handbook supplement. Environmental and Molecular Mutagenesis, 56(9), 724–750. https://doi.org/10.1002/em.21954