
This Key Event Relationship is licensed under the Creative Commons BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
Relationship: 2828
Title
Decrease, AR activation leads to Hypospadias
Upstream event
Downstream event
Key Event Relationship Overview
AOPs Referencing Relationship
| AOP Name | Adjacency | Weight of Evidence | Quantitative Understanding | Point of Contact | Author Status | OECD Status |
|---|---|---|---|---|---|---|
| Androgen receptor (AR) antagonism leading to hypospadias in male (mammalian) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite | ||
| Decreased testosterone synthesis leading to hypospadias in male (mammalian) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite | ||
| 5α-reductase inhibition leading to hypospadias in male (mammalian) offspring | non-adjacent | High | Terje Svingen (send email) | Under development: Not open for comment. Do not cite |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Life Stage Applicability
| Term | Evidence |
|---|---|
| Foetal | High |
Key Event Relationship Description
This non-adjacent KER describes a fetal decrease in androgen receptor (AR) activation in the genital tubercle causing hypospadias in male offspring, postnatally. During fetal development, androgens induce differentiation of the bipotential genital tubercle to a penis, including closure of the urethra. Androgens signal through AR and reduced fetal AR activation can therefore disrupt penis differentiation and lead to the genital malformation hypospadias. Reduced AR activation may happen both through reduced ligand availability (testosterone or dihydrotestosterone (DHT)) and by direct antagonism of AR (Amato et al., 2022; Mattiske & Pask, 2021).
The upstream KE ‘decrease, androgen receptor activation’ (KE 1614) refers to the in vivo event of overall reduction in AR activation. In this case, it therefore refers to a reduction in AR activation in the genital tubercle. Currently, decreased AR activation in mammals is only directly measured in vitro and not in vivo. Instead, indirect assessment of this KE may come from assays measuring AR antagonism, 5α-reductase activity (the enzyme converting testosterone to DHT), or decreased androgen levels (Draskau et al., 2024).
Evidence Collection Strategy
A systematic approach (fig. 1, 2d041pib2_KEr_2828_Figure_1.png (1377×385)) was used to collect evidence based on the methodology described in (Holmer et al., 2024).
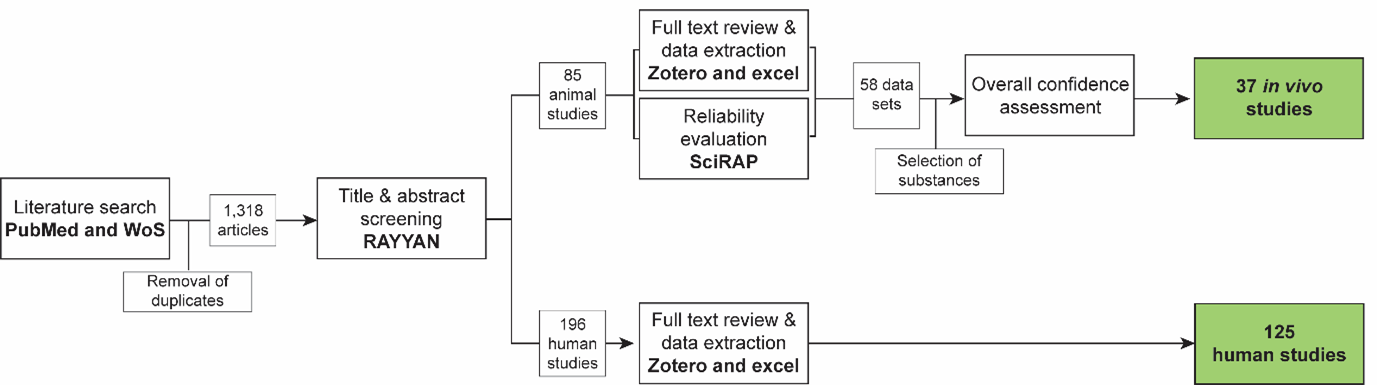{kind=link}
Search strings were synthesized for PubMed and Web of Science Core Collection based on the review question ‘Does decreased androgen receptor activity during fetal development lead to hypospadias in male mammals?’
Search string in PubMed:
("androgen receptor*" OR "testosterone receptor*" OR "receptors, androgen"[MeSH Terms] OR "androgen*" OR "testosterone*" OR "dihydrotestosterone*" OR "androgens"[MeSH Terms] OR "androgen antagonists"[MeSH Terms]) AND ("genital malformation*" OR "hypospadias"[MeSH terms] OR “hypospadia*”).
Search string in Web of Science Core Collection:
TS=(("androgen receptor*" OR "testosterone receptor*" OR "androgen*" OR "testosterone*" OR "dihydrotestosterone*") AND ("hypospadia*" OR "genital malformation*")).
Title & abstract screening:
Retrieved articles were screened in the online tool RAYYAN https://www.rayyan.ai/. After removal of duplicates, the titles and abstracts of the remaining 1,318 articles were screened according to pre-defined exclusion criteria:
- Not English language
- Full text not available
- Not primary literature
- No information about hypospadias in males
- Hypospadias was not coupled to reduced AR activity (measured in vitro, by testosterone levels, mutations, known or suspected anti-androgenic exposure)
During this screening, included papers were split into animal studies and human case studies, respectively.
Full text review, data extraction and reliability evaluation of animal studies:
For animal studies, the full text papers were reviewed using the same exclusion criteria as in the title & abstract screening, and data were extracted from the included papers into an Excel template. In parallel, methodological reliability was assessed using the online tool Science in Risk Assessment and Policy (SciRAP; http://www.scirap.org, see Appendix 1, (56gumbartz_KER_2828__appendix_1.pdf). Based on the SciRAP evaluations, the animal studies were assigned a reliability category using the principles outlined in Table 1. Studies were divided into different datasets, if exposure scenarios led to assignment to different reliability categories. Only studies in reliability categories 1 (reliable without restriction) and 2 (reliable with restriction) were used for the assessment of overall confidence in the data. The animal studies were grouped according to the model substances used. Model substances were selected to include different upstream effects that are known to reduce AR activity: Flutamide, dibutyl phthalate (DBP), vinclozolin, di(2-ethylhexyl) phthalate (DEHP), procymidone, and finasteride.
For each chemical, the overall confidence in the collected data was assessed according to the principles outlined in Table 2.
Table 1 Principles for translation SciRAP evaluations into reliability categories.
|
Reliability Category |
Principles for Categorization |
|
1.Reliable without restriction |
SciRAP methodological quality Score > 80 and all key criteriaa are “Fulfilled” and there are no deficiencies in the non-key criteria that might affect study reliability. |
|
2. Reliable with restriction |
SciRAP methodological quality Score > 65 and one or several of the key criteria are “Partially Fulfilled” or there are minor deficiencies in the non-key criteria that might affect study reliability. |
|
3. Not reliable |
SciRAP methodological quality Score < 65 or one or several of the key criteria are “Not Fulfilled” or there are major deficiencies in the non-key criteria that affect study reliability. |
|
4. Not assignable |
Two or more of the key criteria are “Not Determined” |
aKey criteria were criteria judged as specifically critical for reliability of the data for this KER and were determined a priori. The key criteria for this data collection are outlined in appendix 1, 56gumbartz_KER_2828__appendix_1.pdf.
Table 2 Principles for evaluation of overall confidence in data for each selected substance.
|
Level of confidence |
Principles for Categorizationa |
|
Strong |
OR
|
|
Moderate |
|
|
Weak |
OR
|
|
No effect |
|
a Conflicting results from studies judged as not reliable do not impact categorization.
Full text review and data extraction of human studies
Studies in humans were regarded as supporting evidence, and study quality was not systematically evaluated. All studies were assessed for any major errors in study design or interpretation. The full text papers were reviewed using the same exclusion criteria as in the title & abstract screening, and data were extracted from the included papers into an Excel template. For these studies, the occurrence of hypospadias and the underlying cause were the main data points extracted. The papers were then grouped according to the underlying condition of the reported patients. Four main groups of upstream effects were reported, which were then split into more specific sub-groups:
Effects on AR: This included mutations in AR, extended CAG repeat length in AR, which is associated with reduced AR activity (Chamberlain et al., 1994), and studies in which low AR activity was measured in vitro in genital skin fibroblasts from hypospadias patients.
Effects on 5α-reductase activity: This included mutations in SRD5A2, high testosterone/DHT ratio, and studies in which low 5α-reductase activity was measured in vitro in genital skin fibroblasts from hypospadias patients.
Effects on upstream steroidogenesis: This included mutations in steroidogenesis enzymes (HSD17B3, HSD3B2, or CYP17A1) and patients diagnosed with deficiency in these enzymes based on their hormone and metabolite profile.
Other upstream effects: These were studies, in which low testosterone (basal or hCG-stimulated) was measured in patients, either idiopathic or due to gonadal dysfunction or rare mutations.
The final list of studies was based on an assessment of whether there was a clear description of conditions with decreased AR activity (as described above) and associated hypospadias. Moreover, epidemiologic and case-control studies were included as supporting evidence, and these were included, even if they did not show a causal relationship between reduced AR activity and hypospadias.
Exploration of taxonomic domain of applicability
A separate literature search was performed in “Web of Science, All Databases” to explore the taxonomic domain of applicability of the KER in relation to wildlife species and non-mammalian vertebrates. These studies were not systematically evaluated for reliability but served as supporting evidence for the taxonomic applicability domain of the KERs.
Search string in Web of Science all databases:
hypospadia*and (wildlife OR “animal model” OR domestic OR evolution*).
Evidence Supporting this KER
Biological Plausibility
The biological plausibility for this KER is judged as high. This is largely based on canonical knowledge on normal reproductive development.
The penis originates from a sexually bipotential structure, the genital tubercle, which may differentiate to either a penis or a clitoris, depending on internal cues during fetal development. In males, the fetal testes produce large amounts of testosterone, which can subsequently be converted to the more potent androgen DHT by 5α-reductase in peripheral tissues. Testosterone and DHT both signal through AR in target tissues to initiate masculinization (Amato et al., 2022; Murashima et al., 2015). The critical developmental window for androgen programming of masculinization has been identified in rats as gestational days (GD) 16-20, and is proposed to be gestational weeks 8-14 in humans (Sharpe, 2020; Welsh et al., 2008). As part of the masculinization process orchestrated by androgens, the genital tubercle, which at this point expresses AR in both humans and rodents, differentiates to a penis (Amato & Yao, 2021; Baskin et al., 2020). This includes androgen-mediated elongation of the tubercle, formation of the prepuce, and tubular internalization of urethra, which is closed at the distal tip of the glans penis (Amato et al., 2022). Failure of full closure of the urethra can result in hypospadias, in which the urethra terminates at the ventral side of the penis instead of at the tip (Baskin & Ebbers, 2006; Cohn, 2011).
The dependency of androgens for penile development has been demonstrated in mice with conditional or full knockout of Ar, which results in partly or full sex-reversal of males, including a female-like urethral opening (Willingham et al., 2006; Yucel et al., 2004; Zheng et al., 2015). Similarly, female rats and mice exposed in utero to testosterone present with varying degrees of intersexuality, including, in some cases, a penis (Greene & Ivy, 1937; Zheng et al., 2015).
Empirical Evidence
The empirical evidence for this KER is generally judged as high. This includes evidence from in vivo animal studies and supporting evidence from studies in humans. The upstream KE ‘Decreased AR activation’ refers to an in vivo effect, for which no methods for measurement of this in vivo in mammals currently exist. The effects on the upstream KE were therefore indirectly informed as described in each section.
Animal studies
Effects on the upstream KE were indirectly informed by including animal studies with stressors that are known to reduce AR activity by antagonizing the AR, lowering testosterone production, or inhibiting 5α-reductase. Six stressors, with established anti-androgenic effects, were included (more detailed evaluation of these chemicals can be found in KER-2820 (Holmer et al., 2024)). Table 3 summarizes the empirical evidence and confidence level for each chemical. Details on included evidence is presented in Table 1 in Appendix 2, 9prbqyba2x_Appendix_2_KER_2828.pdf. In summary, all six substances were shown to cause hypospadias in male offspring, and the confidence level for all substances was judged as strong, as conflicting results could be explained (see the section ‘Uncertainties and inconsistencies’). Thus, antagonism or AR, inhibition of 5α-reductase, or reduction in testosterone synthesis, all lead to hypospadias.
Table 3 Summary of empirical evidence for the KER – animal studies. See Table 1 in Appendix 2 ( 9prbqyba2x_Appendix_2_KER_2828.pdf) for details.
|
Chemical |
Upstream effect |
Downstream effect |
Overall confidence |
|
Flutamide |
Androgen receptor antagonist (Simard et al., 1986). |
In utero exposure causes hypospadias in rat and mouse |
Strong |
|
Dibutyl phthalate (DBP) |
Has been shown to reduce fetal intratesticular testosterone and serum testosterone in vivo, but exact mechanism is unknown (Foster, 2006). |
In utero exposure causes hypospadias in rat |
Strong |
|
Vinclozolin |
AR antagonist (Kelce et al., 1994, 1997) |
In utero exposure causes hypospadias in rat and mouse |
Strong |
|
Di(2-ethylhexyl) phthalate (DEHP) |
Has been shown to reduce fetal intratesticular testosterone and serum testosterone in vivo, but exact mechanism is unknown (Parks et al., 2000). |
In utero exposure causes hypospadias in rat |
Strong |
|
Procymidone |
AR antagonist (Ostby et al., 1999). |
In utero exposure causes hypospadias in rat |
Strong |
|
Finasteride |
5α-reductase inhibitor, causing a reduction in DHT (Rittmaster & Wood, 1994). |
In utero exposure causes hypospadias in rat |
Strong |
Supporting human evidence
Effects on the upstream KE were indirectly informed by including studies in humans with a condition (genetic or other) that would reduce or disrupt either 1) function of AR, 2) conversion of testosterone to DHT by disrupting 5α-reductase activity, or 3) production of androgen hormones. Studies measuring low testosterone levels with no underlying cause were also included (see evidence collection strategy). Table 4 lists the studies, in which these conditions were linked to hypospadias in males.
Table 4 Supporting evidence for the KER – human studies. The table lists the number of human studies reporting hypospadias in association with an upstream defect in AR activity, grouped according to the precise effect, and how it was diagnosed (mutation, in vitro activity, or blood hormone and metabolite profile). SRD5A2: 5α-reductase 2; HSD17B3: 17β-hydroxysteroid dehydrogenase 3; HSD3B2: 3β-hydroxysteroid dehydrogenase 2; CYP17A1: 17α-hydroxylase. See Table 2 in Appendix 2 (9prbqyba2x_Appendix_2_KER_2828.pdf) for all included references.
|
Effect on upstream KE |
Supporting studies |
|
Effects on Androgen receptor |
|
|
AR mutations |
27 studies |
|
Extended CAG repeat length in AR |
4 studies |
|
Reduced AR activity (e.g. low receptor binding) in in vitro genital skin fibroblasts |
11 studies |
|
Effects on 5α-reductase activity |
|
|
SRD5A2 mutations |
30 studies |
|
SRD5A2 deficiency, diagnosed by T/DHT-ratio and/or reduced in vitro 5α-reductase activity in genital skin fibroblasts |
8 studies |
|
Effects on upstream steroidogenesis enzymes |
|
|
HSD17B3 mutations |
6 studies |
|
HSD3B2 mutations |
5 studies |
|
CYP17A1 mutation |
1 study |
|
HSD17B3 deficiency, diagnosed by hormone and metabolite profile |
2 studies |
|
HSD3B2 deficiency, diagnosed by hormone and metabolite profile |
4 studies |
|
CYP17A1 deficiency, diagnosed by hormone and metabolite profile |
5 studies |
|
Other upstream effects on low testosterone |
|
|
Low testosterone due to gonadal dysgenesis or hypogonadism |
7 studies |
|
Low basal testosterone or low testosterone response to hCG stimulation. Idiopathic or rare mutations. |
7 studies |
Six case-control studies were extracted, all of which found a correlation between lower testosterone levels (basal or hCG-stimulated) and hypospadias (Austin et al., 2002; Okuyama et al., 1981; Raboch et al., 1976; Ratan et al., 2012; Svensson et al., 1979; Yadav et al., 2011). In two of these studies, the correlation was age-dependent (Austin et al., 2002; Raboch et al., 1976)
One epidemiologic study was extracted, which investigated the association between phthalate exposure and hypospadias risk. Western Australian women exposed through their occupation to phthalates were more likely to have sons with hypospadias (Nassar et al., 2010). It should be noted that there are reported species differences in the effects of phthalates (including DEHP and DBP) on fetal testosterone production between humans, mice, and rats, and the direct translatability of the in vivo evidence is uncertain (Sharpe, 2020).
Dose concordance
Information about dose concordance is not available because AR activity currently cannot be measured in vivo.
Temporal concordance
Direct information about temporal concordance is not available because AR activity currently cannot be measured in vivo.
Indirect information on temporal concordance can be obtained from empirical evidence. In two studies, in which rats were exposed in utero to 750 mg/kg bw/day DBP, intratesticular testosterone levels were reduced in fetal testes, while hypospadias was identified in adult males. Plasma levels of testosterone were also measured in adults, and testosterone levels in exposed males were not significantly different from control males (van den Driesche et al., 2017, 2020). This has also been shown in a study with 500 mg/kg bw/day DBP (Drake et al., 2009). These studies indicate temporal concordance.
Incidence Concordance
Information about dose concordance is not available because AR activity currently cannot be measured in vivo.
Uncertainties and Inconsistencies
The in vivo studies do not directly inform about the upstream KE, ‘decrease, AR activation’. The direct concordance between the KEs can therefore not be determined from the evidence.
For flutamide, two studies reported 100% hypospadias frequencies at doses of 6.25 and 10 mg/kg bw/day (Goto et al., 2004; McIntyre et al., 2001), while another study found a frequency of 56.9% when giving 20 mg/kg bw/day (Kita et al., 2016). This might be explained by a longer exposure window in the first two studies and uncertainties in assessment of hypospadias.
For DBP, there were discrepancies in whether 250 mg/kg bw/day was LOAEL (Mylchreest et al., 1998, 1999) or NOAEL (Jiang et al., 2007) for DBP. This conflict was explained by differences in exposure windows, supported by the observation that the frequency of hypospadias at 250 mg/kg bw/day was reported as very low (Mylchreest et al., 1998, 1999).
One study with vinclozolin (Ostby J et al., 1999) and one with procymidone (Hass et al., 2012) did not find hypospadias after in utero exposure. In both cases, this was likely due to too low doses tested.
In most of the human studies of steroidogenesis deficiency, serum or plasma levels of testosterone were reduced at baseline and/or upon hCG stimulation (Al-Sinani et al., 2015; Ammini et al., 1997; Cara et al., 1985; Chen, Huang, et al., 2021; Dean et al., 1984; Galli-Tsinopoulou et al., 2018; Imperato-McGinley et al., 1979; Kaufman et al., 1983; Mendonca et al., 1987, 2000; Neocleous et al., 2012; New, 1970; Pang et al., 1983; Perrone et al., 1985; Rabbani et al., 2012; Sherbet et al., 2003), but in a few studies, testosterone levels were normal (Donadille et al., 2018; Kon et al., 2015; Luna et al., 2021). In these cases, the effect of these deficiencies on tissue AR activitation is uncertain.
For AR CAG repeat length, a case-control study did not find an association with hypospadias (Radpour R et al., 2007), but this could be because the hypospadias cases included had other etiologies.
Lastly, as there are currently no universal guidelines for identification and scoring of hypospadias in rodents, there are large variations in methods of assessment, and minor cases of hypospadias may be overlooked in some studies and included in others. This poses an uncertainty in the frequency reports in the scientific evidence.
Known modulating factors
|
Modulating Factors |
MF details |
Effects on the KER |
References |
|
AR CAG repeat length |
Extended CAG repeat length in AR is associated with reduced AR activity |
Higher risk of hypospadias development |
(Chamberlain et al., 1994) |
Quantitative Understanding of the Linkage
The quantitative understanding of the relationship is low. As there are currently no direct measurement methods of the upstream KE (reduced AR activation) in mammals, quantification of the relationship is difficult to assess.
Response-response Relationship
A model for phthalates has been developed, aiming to predict the frequency of hypospadias in male offspring based on reductions in ex vivo testosterone production, an indirect indication of AR activation. In this model, hypospadias was induced from around a 60% reduction in testosterone levels. The model does not consider hypospadias severity and is only for phthalate chemicals (Earl Gray et al., 2024).
Time-scale
The time-scale of this KER depends on the species but is likely days to weeks.
AR activation happens within minutes, from ligand binding to nuclear translocation and promotor activation (Nightingale et al., 2003; Schaufele et al., 2005), while transcriptional and translational effects are observed minutes to hours later (Kang et al., 2002). AR programming of the genital tubercle occurs during fetal development in the masculinization programming window (Sharpe, 2020). The time-scale for morphological effects in the tissue then depends on the species. In humans, penis development is completed prior to birth and hypospadias can be observed at birth. In rodents, penis development is not fully completed until weeks after birth, but hypospadias can often be observed earlier than this (Table 3).
Known Feedforward/Feedback loops influencing this KER
Local disruption of AR activation in the genital tubercle irreversibly disrupts development, so there are no known feedforward/feedback loops.
Domain of Applicability
Taxonomic applicability
In mammals, androgens are one of the primary drivers of penis differentiation. Hypospadias has been observed in several mammals, but most frequently reported in laboratory rodents and in humans (Chang et al., 2020; S. Wang & Zheng, 2025). In vivo studies in rats and mice show that in utero exposure to anti-androgenic chemicals can cause hypospadias in male offspring (see Table 3). Many human case studies report boys born with hypospadias and associated deficiency in steroid hormone synthesis, 5α-reductase activity, or AR activity (see table 4).
The biologically plausible domain of applicability may extend beyond the empirical domain because androgen-controlled development of male external genitalia is evolutionary conserved in most mammals and, to some extent, also in other vertebrate classes (Gredler et al., 2014). Hypospadias can in principle occur in all animals that form a genital tubercle and have been observed in many domestic animal species including dog (Sonne et al., 2008; Switonski et al., 2018), cat (Nowacka-Woszuk et al., 2014), cattle (Murakami, 2008), sheep (Smith et al., 2012), and horse (De Lorenzi et al., 2010) as well as in wildlife species such as polar bear (Stamper et al., 1999), giraffe (Meuffels et al., 2020), and Tamar Wallaby (Leihy et al., 2011). The observed hypospadias in these animals is not, per se, linked to anti-androgenic exposure, which has only been sparsely investigated in other species than mice, rats, and humans. One study in monkeys did show hypospadias upon oral exposure to finasteride (Prahalada et al., 1997), and bicalutamide exposure induced hypospadias in guinea pigs (S. Wang et al., 2018). A study in rabbits exposed to procymidone did not find hypospadias in males (Inawaka et al., 2010). Another study in hyenas did also not find hypospadias in males after exposure to the anti-androgen finasteride (Drea et al., 1998), but it should be noted that the hyenas have a remarkable sexual development where penile growth occur in both females and males before androgen synthesis is initiated (Cunha et al., 2014) (the studies in hyena and rabbit were identified in our evidence collection but were judged as ‘unreliable’ and therefore not included as empirical evidence).
Sex applicability
The AR is expressed in the fetal genital tubercle of both females and males (Amato & Yao, 2021; Baskin et al., 2020), but hypospadias is primarily a term used for a malformation of the penis (Baskin & Ebbers, 2006), limiting the applicability of this KER to males.
Life stage applicability
Differentiation of the penis occurs during fetal life in the masculinization programming window (GD 16-20 in rats, around GW 8-14 in humans), when androgen production is high (Welsh et al., 2008; C. Wolf et al., 2000a). In rats, exposure to anti-androgenic chemicals outside of, or in the late part of the masculinization programming window does not cause hypospadias or only to a low degree (Clark et al., 1993; van den Driesche et al., 2017; C. Wolf et al., 2000a), while exposure in the earlier (or full) window causes a higher frequency of hypospadias (depending on dose and chemical) (Table 3). In humans, hypospadias can be diagnosed at birth (X. Yu et al., 2019), while in rodents, some parts of penis development occur postnatally (Schlomer et al., 2013; Sinclair et al., 2017). In these species, hypospadias may be observed at birth but is optimally diagnosed and severity classified weeks later. Given that disruptions to androgen programming takes place in fetal life, even though the AO is best detected postnatally, the life stage applicability is defined as fetal life.
References
Al-Sinani, A., Mula-Abed, W., Al-Kindi, M., Al-Kusaibi, G., Al-Azkawi, H., & Nahavandi, N. (2015). A Novel Mutation Causing 17-β-Hydroxysteroid Dehydrogenase Type 3 Deficiency in an Omani Child: First Case Report and Review of Literature. Oman Medical Journal, 30(2), 129–134. https://doi.org/10.5001/omj.2015.27
Amato, C. M., & Yao, H. H.-C. (2021). Developmental and sexual dimorphic atlas of the prenatal mouse external genitalia at the single-cell level. Proceedings of the National Academy of Sciences of the United States of America, 118(25). https://doi.org/10.1073/pnas.2103856118
Amato, C. M., Yao, H. H.-C., & Zhao, F. (2022). One Tool for Many Jobs: Divergent and Conserved Actions of Androgen Signaling in Male Internal Reproductive Tract and External Genitalia. Frontiers in Endocrinology, 13, 910964. https://doi.org/10.3389/fendo.2022.910964
Ammini, A., Sharma, D., Gupta, R., Mohapatra, I., Kucheria, K., Kriplani, A., Takkar, D., Mitra, D., & Vijayaraghavan, M. (1997). Familial male pseudohermaphroditism. Indian Journal of Pediatrics, 64(3), 419–423. https://doi.org/10.1007/BF02845218
Austin, P., Siow, Y., Fallat, M., Cain, M., Rink, R., & Casale, A. (2002). The relationship between müllerian inhibiting substance and androgens in boys with hypospadias. The Journal of Urology, 168(4), 1784–1788; discussion 1788. https://doi.org/10.1097/01.ju.0000023680.64155.5c
Baskin, L., Cao, M., Sinclair, A., Li, Y., Overland, M., Isaacson, D., & Cunha, G. R. (2020). Androgen and estrogen receptor expression in the developing human penis and clitoris. Differentiation; Research in Biological Diversity, 111, 41–59. https://doi.org/10.1016/j.diff.2019.08.005
Baskin, L., & Ebbers, M. (2006). Hypospadias: Anatomy, etiology, and technique. Journal of Pediatric Surgery, 41(3), 463–472. https://doi.org/10.1016/j.jpedsurg.2005.11.059
Cara, J., Jr Moshang, T., Bongiovanni, A., & Marx, B. (1985). Elevated 17-hydroxyprogesterone and testosterone in a newborn with 3-beta-hydroxysteroid dehydrogenase deficiency. The New England Journal of Medicine, 313(10), 618–621. https://doi.org/10.1056/NEJM198509053131007
Chamberlain, N. L., Driver, E. D., & Miesfeld, R. L. (1994). The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactivation function. Nucleic Acids Research, 22(15), 3181–3186. https://doi.org/10.1093/nar/22.15.3181
Chang, J., Wang, S., & Zheng, Z. (2020). Etiology of Hypospadias: A Comparative Review of Genetic Factors and Developmental Processes Between Human and Animal Models. Research and Reports in Urology, Volume 12, 673–686. https://doi.org/10.2147/RRU.S276141
Chen, L., Huang, H., Zhang, H., Zhu, G., & Zhu, M. (2021). Three cases of 3β-hydroxysteroid dehydrogenase deficiency: Clinical analysis. Advances in Clinical and Experimental Medicine : Official Organ Wroclaw Medical University, 30(3), 289–299. https://doi.org/10.17219/acem/131220
Clark, R., Anderson, C., Prahalada, S., Robertson, R., Lochry, E., Leonard, Y., Stevens, J., & Hoberman, A. (1993). Critical developmental periods for effects on male rat genitalia induced by finasteride, a 5 alpha-reductase inhibitor. Toxicology and Applied Pharmacology, 119(1), 34–40. https://doi.org/10.1006/taap.1993.1041
Cohn, M. J. (2011). Development of the external genitalia: Conserved and divergent mechanisms of appendage patterning. Developmental Dynamics, 240(5), 1108–1115. https://doi.org/10.1002/dvdy.22631
Cunha, G. R., Risbridger, G., Wang, H., Place, N. J., Grumbach, M., Cunha, T. J., Weldele, M., Conley, A. J., Barcellos, D., Agarwal, S., Bhargava, A., Drea, C., Hammond, G. L., Siiteri, P., Coscia, E. M., McPhaul, M. J., Baskin, L. S., & Glickman, S. E. (2014). Development of the external genitalia: Perspectives from the spotted hyena (Crocuta crocuta). Differentiation, 87(1–2), 4–22. https://doi.org/10.1016/j.diff.2013.12.003
De Lorenzi, L., Genualdo, V., Iannuzzi, A., Di Meo, G. P., Perucatti, A., Mancuso, R., Russo, M., Di Berardino, D., Parma, P., & Iannuzzi, L. (2010). Cytogenetic and Genetic Studies in a Hypospadic Horse (Equus caballus, 2n = 64). Sexual Development, 4(6), 352–357. https://doi.org/10.1159/000319527
Dean, H., Shackleton, C., & Winter, J. (1984). Diagnosis and natural history of 17-hydroxylase deficiency in a newborn male. The Journal of Clinical Endocrinology and Metabolism, 59(3), 513–520. https://doi.org/10.1210/jcem-59-3-513
Donadille, B., Houang, M., Netchine, I., Siffroi, J., & Christin-Maitre, S. (2018). Human 3beta-hydroxysteroid dehydrogenase deficiency associated with normal spermatic numeration despite a severe enzyme deficit. Endocrine Connections, 7(3), 395–402. https://doi.org/10.1530/EC-17-0306
Drake, A., van den Driesche, S., Scott, H., Hutchison, G., Seckl, J., & Sharpe, R. (2009). Glucocorticoids amplify dibutyl phthalate-induced disruption of testosterone production and male reproductive development. Endocrinology, 150(11), 5055–5064. https://doi.org/10.1210/en.2009-0700
Draskau, M. K., Rosenmai, A. K., Bouftas, N., Johansson, H. K. L., Panagiotou, E. M., Holmer, M. L., Elmelund, E., Zilliacus, J., Beronius, A., Damdimopolou, P., van Duursen, M., & Svingen, T. (2024). AOP Report: An Upstream Network for Reduced Androgen Signaling Leading to Altered Gene Expression of Androgen Receptor-Responsive Genes in Target Tissues. Environmental Toxicology and Chemistry. https://doi.org/10.1002/etc.5972
Drea, C., Weldele, M., Forger, N., Coscia, E., Frank, L., Licht, P., & Glickman, S. (1998). Androgens and masculinization of genitalia in the spotted hyaena (Crocuta crocuta). 2. Effects of prenatal anti-androgens. Journal of Reproduction and Fertility, 113(1), 117–127. https://doi.org/10.1530/jrf.0.1130117
Earl Gray, L. J., Lambright, C., Evans, N., Ford, J., & Conley, M. (2024). Using targeted fetal rat testis genomic and endocrine alterations to predict the effects of a phthalate mixture on the male reproductive tract. Current Research in Toxicology, 7, 100180. https://doi.org/10.1016/j.crtox.2024.100180
Foster, P. M. D. (2006). Disruption of reproductive development in male rat offspring following in utero exposure to phthalate esters. International Journal of Andrology, 29(1), 140–147. https://doi.org/10.1111/j.1365-2605.2005.00563.x
Galli-Tsinopoulou, A., Serbis, A., KotanidouP, E., Litou, E., Dokousli, V., Mouzaki, K., Fanis, P., Neocleous, V., & Skordis, N. (2018). 46,XY Disorder of Sex Development due to 17-Beta Hydroxysteroid Dehydrogenase Type 3 Deficiency in an Infant of Greek Origin. Journal of Clinical Research in Pediatric Endocrinology, 10(1), 74–78. https://doi.org/10.4274/jcrpe.4829
Goto, K., Koizumi, K., Takaori, H., Fujii, Y., Furuyama, Y., Saika, O., Suzuki, H., Saito, K., & Suzuki, K. (2004). Effects of flutamide on sex maturation and behavior of offspring born to female rats treated during late pregnancy. The Journal of Toxicological Sciences, 29(5), 517–534. https://doi.org/10.2131/jts.29.517
Gredler, M. L., Larkins, C. E., Leal, F., Lewis, A. K., Herrera, A. M., Perriton, C. L., Sanger, T. J., & Cohn, M. J. (2014). Evolution of External Genitalia: Insights from Reptilian Development. Sexual Development, 8(5), 311–326. https://doi.org/10.1159/000365771
Greene, R. R., & Ivy, A. C. (1937). THE EXPERIMENTAL PRODUCTION OF INTERSEXUALITY IN THE FEMALE RAT WITH TESTOSTERONE. Science (New York, N.Y.), 86(2226), 200–201. https://doi.org/10.1126/science.86.2226.200-a
Hass, U., Boberg, J., Christiansen, S., Jacobsen, P., Vinggaard, A., Taxvig, C., Poulsen, M., Herrmann, S., Jensen, B., Petersen, A., Clemmensen, L., & Axelstad, M. (2012). Adverse effects on sexual development in rat offspring after low dose exposure to a mixture of endocrine disrupting pesticides. Reproductive Toxicology (Elmsford, N.Y.), 34(2), 261–274. https://doi.org/10.1016/j.reprotox.2012.05.090
Holmer, M. L., Zilliacus, J., Draskau, M. K., Hlisníková, H., Beronius, A., & Svingen, T. (2024). Methodology for developing data-rich Key Event Relationships for Adverse Outcome Pathways exemplified by linking decreased androgen receptor activity with decreased anogenital distance. Reproductive Toxicology, 128, 108662. https://doi.org/10.1016/j.reprotox.2024.108662
Imperato-McGinley, J., Peterson, R., Stoller, R., & Goodwin, W. (1979). Male pseudohermaphroditism secondary to 17 beta-hydroxysteroid dehydrogenase deficiency: Gender role change with puberty. The Journal of Clinical Endocrinology and Metabolism, 49(3), 391–395. https://doi.org/10.1210/jcem-49-3-391
Inawaka, K., Kishimoto, N., Higuchi, H., & Kawamura, S. (2010). Maternal exposure to procymidone has no effects on fetal external genitalia development in male rabbit fetuses in a modified developmental toxicity study. JOURNAL OF TOXICOLOGICAL SCIENCES, 35(3), 299–307. https://doi.org/10.2131/jts.35.299
Jiang, J., Ma, L., Yuan, L., Wang, X., & Zhang, W. (2007). Study on developmental abnormalities in hypospadiac male rats induced by maternal exposure to di-n-butyl phthalate (DBP). Toxicology, 232(3), 286–293. https://doi.org/10.1016/j.tox.2007.01.018
Jiang, J., Zhong, C., Zhu, Y., XuL, D., Wood, K., Sun, W., Li, E., Liu, Z., Zhao, W., Ruan, Y., & Xia, S. (2016). Prenatal exposure to di-n-butyl phthalate (DBP) differentially alters androgen cascade in undeformed versus hypospadiac male rat offspring. Reproductive Toxicology (Elmsford, N.Y.), 61, 75–81. https://doi.org/10.1016/j.reprotox.2016.02.016
Kang, H.-Y., Huang, K.-E., Chang, S. Y., Ma, W.-L., Lin, W.-J., & Chang, C. (2002). Differential Modulation of Androgen Receptor-mediated Transactivation by Smad3 and Tumor Suppressor Smad4. Journal of Biological Chemistry, 277(46), 43749–43756. https://doi.org/10.1074/jbc.M205603200
Kaufman, F., Costin, G., Goebelsmann, U., Stanczyk, F., & Zachmann, M. (1983). Male pseudohermaphroditism due to 17,20-desmolase deficiency. The Journal of Clinical Endocrinology and Metabolism, 57(1), 32–36. https://doi.org/10.1210/jcem-57-1-32
Kelce, W. R., Lambright, C. R., Gray, L. E., & Roberts, K. P. (1997). Vinclozolin andp,p′-DDE Alter Androgen-Dependent Gene Expression:In VivoConfirmation of an Androgen Receptor-Mediated Mechanism. Toxicology and Applied Pharmacology, 142(1), 192–200. https://doi.org/10.1006/taap.1996.7966
Kelce, W. R., Monosson, E., Gamcsik, M. P., Laws, S. C., & Gray, L. E. (1994). Environmental Hormone Disruptors: Evidence That Vinclozolin Developmental Toxicity Is Mediated by Antiandrogenic Metabolites. Toxicology and Applied Pharmacology, 126(2), 276–285. https://doi.org/10.1006/taap.1994.1117
Kita, D., Meyer, K., Venturelli, A., Adams, R., Machado, D., Morais, R., Swan, S., Gennings, C., & Martino-Andrade, A. (2016). Manipulation of pre and postnatal androgen environments and anogenital distance in rats. TOXICOLOGY, 368, 152–161. https://doi.org/10.1016/j.tox.2016.08.021
Kon, M., Suzuki, E., Dung, V., Hasegawa, Y., Mitsui, T., Muroyas, K., Ueoka, K., Igarashi, N., Nagasaki, K., Oto, Y., Hamajima, T., Yoshino, K., Igarashi, M., Kato-Fukui, Y., Nakabayashi, K., Hayashi, K., Hata, K., Matsubara, Y., Moriya, K., … Fukami, M. (2015). Molecular basis of non-syndromic hypospadias: Systematic mutation screening and genome-wide copy-number analysis of 62 patients. HUMAN REPRODUCTION, 30(3), 499–506. https://doi.org/10.1093/humrep/deu364
Leihy, M. W., Shaw, G., Wilson, J. D., & Renfree, M. B. (2011). Development of the Penile Urethra in the Tammar Wallaby. Sexual Development, 5(5), 241–249. https://doi.org/10.1159/000334053
Luna, S., Wegner, D., Gale, S., Yang, P., Hollander, A., St Dennis-Feezle, L., Nabhan, Z., Ory, D., Cole, F., & Wambach, J. (2021). Whole exome sequencing and functional characterization increase diagnostic yield in siblings with a 46, XY difference of sexual development (DSD). The Journal of Steroid Biochemistry and Molecular Biology, 212, 105908. https://doi.org/10.1016/j.jsbmb.2021.105908
Mattiske, D. M., & Pask, A. J. (2021). Endocrine disrupting chemicals in the pathogenesis of hypospadias; developmental and toxicological perspectives. Current Research in Toxicology, 2, 179–191. https://doi.org/10.1016/j.crtox.2021.03.004
McIntyre, B., Barlow, N., & Foster, P. (2001). Androgen-mediated development in male rat offspring exposed to flutamide in utero: Permanence and correlation of early postnatal changes in anogenital distance and nipple retention with malformations in androgen-dependent tissues. Toxicological Sciences : An Official Journal of the Society of Toxicology, 62(2), 236–249. https://doi.org/10.1093/toxsci/62.2.236
Mendonca, B., Bloise, W., Arnhold, I., Batista, M., Toledo, S., Drummond, M., Nicolau, W., & Mattar, E. (1987). Male pseudohermaphroditism due to nonsalt-losing 3 beta-hydroxysteroid dehydrogenase deficiency: Gender role change and absence of gynecomastia at puberty. Journal of Steroid Biochemistry, 28(6), 669–675. https://doi.org/10.1016/0022-4731(87)90396-7
Mendonca, B., Inacio, M., Arnhold, I., Costa, E., Bloise, W., Martin, R., Denes, F., Silva, F., Andersson, S., Lindqvist, A., & Wilson, J. (2000). Male pseudohermaphroditism due to 17 beta-hydroxysteroid dehydrogenase 3 deficiency. Diagnosis, psychological evaluation, and management. Medicine, 79(5), 299–309. https://doi.org/10.1097/00005792-200009000-00003
Meuffels, J., Luther-Binoir, I., Daffue, W., Deacon, F., & Mitchell, E. P. (2020). Testicular disorder of sexual development with cryptorchidism, penile hypoplasia and hypospadias in a giraffe (Giraffa camelopardalis giraffa). JOURNAL OF THE SOUTH AFRICAN VETERINARY ASSOCIATION, 91. https://doi.org/10.4102/jsava.v91i0.1971
Murakami, T. (2008). Anatomical Examination of Hypospadias in Cattle. Journal of the Japan Veterinary Medical Association, 61(12), 931–935. https://doi.org/10.12935/jvma1951.61.931
Murashima, A., Kishigami, S., Thomson, A., & Yamada, G. (2015). Androgens and mammalian male reproductive tract development. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms, 1849(2), 163–170. https://doi.org/10.1016/j.bbagrm.2014.05.020
Mylchreest, E., Cattley, R., & Foster, P. (1998). Male reproductive tract malformations in rats following gestational and lactational exposure to Di(n-butyl) phthalate: An antiandrogenic mechanism? Toxicological Sciences : An Official Journal of the Society of Toxicology, 43(1), 47–60. https://doi.org/10.1006/toxs.1998.2436
Mylchreest, E., Sar, M., Cattley, R., & Foster, P. (1999). Disruption of androgen-regulated male reproductive development by di(n-butyl) phthalate during late gestation in rats is different from flutamide. Toxicology and Applied Pharmacology, 156(2), 81–95. https://doi.org/10.1006/taap.1999.8643
Nassar, N., Abeywardana, P., Barker, A., & Bower, C. (2010). Parental occupational exposure to potential endocrine disrupting chemicals and risk of hypospadias in infants. Occupational and Environmental Medicine, 67(9), 585–589. https://doi.org/10.1136/oem.2009.048272
Neocleous, V., Sismani, C., Shammas, C., Efstathiou, E., Alexandrou, A., Ioannides, M., Argyrou, M., Patsalis, P., Phylactou, L., & Skordis, N. (2012). Duplication of exons 3-10 of the HSD17B3 gene: A novel type of genetic defect underlying 17 beta-HSD-3 deficiency. GENE, 499(2), 250–255. https://doi.org/10.1016/j.gene.2012.03.031
New, M. (1970). Male pseudohermaphroditism due to 17 alpha-hydroxylase deficiency. The Journal of Clinical Investigation, 49(10), 1930–1941. https://doi.org/10.1172/JCI106412
Nightingale, J., Chaudhary, K. S., Abel, P. D., Stubbs, A. P., Romanska, H. M., Mitchell, S. E., Stamp, G. W. H., & Lalani, E.-N. (2003). Ligand Activation of the Androgen Receptor Downregulates E-Cadherin-Mediated Cell Adhesion and Promotes Apoptosis of Prostatic Cancer Cells. Neoplasia, 5(4), 347–361. https://doi.org/10.1016/S1476-5586(03)80028-3
Nowacka-Woszuk, J., Szczerbal, I., Salamon, S., Kociucka, B., Jackowiak, H., Prozorowska, E., Slaska, B., Rozanska, D., Orzelski, M., Ochota, M., Dzimira, S., Lipiec, M., Nizanski, W., & Switonski, M. (2014). Testicular disorder of sex development in four cats with a male karyotype (38,XY; SRY-positive). Animal Reproduction Science, 151(1–2), 42–48. https://doi.org/10.1016/j.anireprosci.2014.10.001
Okuyama, A., Namiki, M., Koide, T., Itatani, H., Nishimoto, N., Mizutani, S., Sonoda, T., Aono, T., & Matsumoto, K. (1981). Pituitary and gonadal function in prepubertal and pubertal boys with hypospadias. Acta Endocrinologica, 98(3), 464–469. https://doi.org/10.1530/acta.0.0980464
Ostby, J., Kelce, W. R., Lambright, C., Wolf, C. J., Mann, P., & Gray, L. E. (1999). The fungicide procymidone alters sexual differentiation in the male rat by acting as an androgen-receptor antagonist in vivo and in vitro. Toxicology and Industrial Health, 15(1–2), 80–93. https://doi.org/10.1177/074823379901500108
Ostby J, Monosson E, Kelce WR, & Gray, L. J. (1999). Environmental antiandrogens: Low doses of the fungicide vinclozolin alter sexual differentiation of the male rat. Toxicology and Industrial Health, 15(1), 48–64. https://doi.org/10.1177/074823379901500106
Pang, S., Levine, L., Stoner, E., Opitz, J., Pollack, M., Dupont, B., & New, M. (1983). Nonsalt-losing congenital adrenal hyperplasia due to 3 beta-hydroxysteroid dehydrogenase deficiency with normal glomerulosa function. The Journal of Clinical Endocrinology and Metabolism, 56(4), 808–818. https://doi.org/10.1210/jcem-56-4-808
Parks, L. G., Ostby, J. S., Lambright, C. R., Abbott, B.D., Klinefelter, G. R., Barlow, N. J., & Gray LE Jr. (2000). The Plasticizer Diethylhexyl Phthalate Induces Malformations by Decreasing Fetal Testosterone Synthesis during Sexual Differentiation in the Male Rat. Toxicological Sciences, 58(2), 339–349. https://doi.org/10.1093/toxsci/58.2.339
Perrone, L., Criscuolo, T., Sinisi, A., Graziani, M., Manzo, T., Sicuranza, R., Bellastella, A., & Faggiano, M. (1985). Male pseudohermaphroditism due to 3 beta-hydroxysteroid dehydrogenase-isomerase deficiency associated with atrial septal defect. Acta Endocrinologica, 110(4), 532–539. https://doi.org/10.1530/acta.0.1100532
Prahalada, S., Tarantal, A., Harris, G., Ellsworth, K., Clarke, A., Skiles, G., MacKenzie, K., Kruk, L., Ablin, D., Cukierski, M., Peter, C., vanZwieten, M., & Hendrickx, A. (1997). Effects of finasteride, a type 2 5-alpha reductase inhibitor, on fetal development in the rhesus monkey (Macaca mulatta). Teratology, 55(2), 119–131. https://doi.org/10.1002/(SICI)1096-9926(199702)55:2%253C119::AID-TERA1%253E3.0.CO;2-Z
Rabbani, B., Mahdieh, N., Ashtiani, M. H., Setoodeh, A., & A Rabbani. (2012). In silico structural, functional and pathogenicity evaluation of a novel mutation: An overview of HSD3B2 gene mutations. Gene, 503(2), 215–221. https://doi.org/10.1016/j.gene.2012.04.080
Raboch, J., Pondelickova, J., & Starka, L. (1976). Plasma testosterone values in hypopspadiacs. Andrologia, 8(3), 255–258. https://doi.org/10.1111/j.1439-0272.1976.tb02144.x
Radpour R, Rezaee M, Tavasoly A, Solati S, & Saleki A. (2007). Association of long polyglycine tracts (GGN repeats) in exon 1 of the androgen receptor gene with cryptorchidism and penile hypospadias in Iranian patients. Journal of Andrology, 28(1), 164–169. https://doi.org/10.2164/jandrol.106.000927
Ratan, S., Aggarwal, S., Mishra, T., Saxena, A., Yadav, S., Pandey, R., Sharma, A., & Dhanwal, D. (2012). Children with isolated hypospadias have different hormonal profile compared to those with associated anomalies. Journal of Pediatric Endocrinology & Metabolism : JPEM, 25(1), 111–119. https://doi.org/10.1515/jpem.2011.421
Rittmaster, R. S., & Wood, A. J. J. (1994). Finasteride. New England Journal of Medicine, 330(2), 120–125. https://doi.org/10.1056/NEJM199401133300208
Schaufele, F., Carbonell, X., Guerbadot, M., Borngraeber, S., Chapman, M. S., Ma, A. A. K., Miner, J. N., & Diamond, M. I. (2005). The structural basis of androgen receptor activation: Intramolecular and intermolecular amino–carboxy interactions. Proceedings of the National Academy of Sciences, 102(28), 9802–9807. https://doi.org/10.1073/pnas.0408819102
Schlomer, B. J., Feretti, M., Rodriguez, E. J., Blaschko, S., Cunha, G., & Baskin, L. (2013). Sexual differentiation in the male and female mouse from days 0 to 21: A detailed and novel morphometric description. The Journal of Urology, 190(4 Suppl), 1610–1617. https://doi.org/10.1016/j.juro.2013.02.3198
Sharpe, R. M. (2020). Androgens and the masculinization programming window: Human-rodent differences. Biochemical Society Transactions, 48(4), 1725–1735. https://doi.org/10.1042/BST20200200
Sherbet, D., Tiosano, D., Kwist, K., Hochberg, Z., & RJ Auchus. (2003). CYP17 mutation E305G causes isolated 17,20-lyase deficiency by selectively altering substrate binding. The Journal of Biological Chemistry, 278(49), 48563–48569. https://doi.org/10.1074/jbc.M307586200
Simard, J., Luthy, I., Guay, J., Bélanger, A., & Labrie, F. (1986). Characteristics of interaction of the antiandrogen flutamide with the androgen receptor in various target tissues. Molecular and Cellular Endocrinology, 44(3), 261–270. https://doi.org/10.1016/0303-7207(86)90132-2
Sinclair, A., Cao, M., Pask, A., Baskin, L., & Cunha, G. (2017). Flutamide-induced hypospadias in rats: A critical assessment. Differentiation; Research in Biological Diversity, 94, 37–57. https://doi.org/10.1016/j.diff.2016.12.001
Smith, K., Brown, P., & Barr, F. (2012). A Survey of Congenital Reproductive Abnormalities in Rams in Abattoirs in South West England. Reproduction in Domestic Animals, 47(5), 740–745. https://doi.org/10.1111/j.1439-0531.2011.01952.x
Sonne, C., Dietz, R., Born, E. W., Leifsson, P. S., & Andersen, S. (2008). Is there a link between hypospadias and organochlorine exposure in East Greenland sledge dogs (Canis familiaris)? Ecotoxicology and Environmental Safety, 69(3), 391–395. https://doi.org/10.1016/j.ecoenv.2007.09.008
Stamper, M. A., Norton, T., Spodnick, G., Marti, J., & Loomis, M. (1999). Hypospadias in a polar bear (Ursus maritimus). Journal of Zoo and Wildlife Medicine: Official Publication of the American Association of Zoo Veterinarians, 30(1), 141–144.
Svensson, J., Eneroth, P., Gustafsson, J., Ritzén, M., & Stenberg, A. (1979). Reduction of androstenedione by skin in vitro and serum levels of gonadotrophins and androgens in men with hypospadias. The Journal of Endocrinology, 82(3), 395–401. https://doi.org/10.1677/joe.0.0820395
Switonski, M., Dzimira, S., Aleksiewicz, R., Szczerbal, I., Nowacka-Woszuk, J., Krzeminska, P., Deska, T., & Nizanski, W. (2018). Hypospadias Is Not Rare in Dogs: Five New Cases, a Retrospective Study, and a Review of the Literature. Sexual Development : Genetics, Molecular Biology, Evolution, Endocrinology, Embryology, and Pathology of Sex Determination and Differentiation, 12(5), 244–250. https://doi.org/10.1159/000490079
van den Driesche, S., Kilcoyne, K., Wagner, I., Rebourcet, D., Boyle, A., Mitchell, R., McKinnell, C., Macpherson, S., Donat, R., Shukla, C., Jorgensen, A., Meyts, E., Skakkebaek, N., & Sharpe, R. (2017). Experimentally induced testicular dysgenesis syndrome originates in the masculinization programming window. JCI Insight, 2(6), e91204. https://doi.org/10.1172/jci.insight.91204
van den Driesche, S., Shoker, S., Inglis, F., Palermo, C., Langsch, A., & Otter, R. (2020). Systematic comparison of the male reproductive tract in fetal and adult Wistar rats exposed to DBP and DINP in utero during the masculinisation programming window. Toxicology Letters, 335, 37–50. https://doi.org/10.1016/j.toxlet.2020.10.006
Wang, S., Shi, M., Zhu, D., Mathews, R., & Zheng, Z. (2018). External Genital Development, Urethra Formation, and Hypospadias Induction in Guinea Pig: A Double Zipper Model for Human Urethral Development. Urology, 113, 179–186. https://doi.org/10.1016/j.urology.2017.11.002
Wang, S., & Zheng, Z. (2025). Differences in Formation of Prepuce and Urethral Groove During Penile Development Between Guinea Pigs and Mice Are Controlled by Differential Expression of Shh, Fgf10 and Fgfr2. Cells, 14(5), 348. https://doi.org/10.3390/cells14050348
Welsh, M., Saunders, P., Fisken, M., Scott, H., Hutchison, G., Smith, L., & Sharpe, R. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. The Journal of Clinical Investigation, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241
Willingham, E., Agras, K., Souza, A. J. de, Konijeti, R., Yucel, S., Rickie, W., Cunha, G., & Baskin, L. (2006). Steroid receptors and mammalian penile development: An unexpected role for progesterone receptor? The Journal of Urology, 176(2), 728–733. https://doi.org/10.1016/j.juro.2006.03.078
Wolf, C., LeBlanc, G., Ostby, J., & Gray, L. J. (2000). Characterization of the period of sensitivity of fetal male sexual development to vinclozolin. Toxicological Sciences : An Official Journal of the Society of Toxicology, 55(1), 152–161. https://doi.org/10.1093/toxsci/55.1.152
Yadav, C., Bajpai, M., Kumar, V., Datta, S., Gupta, P., Ahmed, R., & Banerjee, B. (2011). Polymorphisms in the P450 c17 (17-hydroxylase/17, 20-Lyase) gene: Association with estradiol and testosterone concentration in hypospadias. Urology, 78(4), 902–907. https://doi.org/10.1016/j.urology.2011.04.021
Yu, X., Nassar, N., Mastroiacovo, P., Canfield, M., Groisman, B., Bermejo-Sánchez, E., Ritvanen, A., Kiuru-Kuhlefelt, S., Benavides, A., Sipek, A., Pierini, A., Bianchi, F., Källén, K., Gatt, M., Morgan, M., Tucker, D., Canessa, M. A., Gajardo, R., Mutchinick, O. M., … Agopian, A. J. (2019). Hypospadias Prevalence and Trends in International Birth Defect Surveillance Systems, 1980-2010. European Urology, 76(4), 482–490. https://doi.org/10.1016/j.eururo.2019.06.027
Yucel, S., Liu, W., Cordero, D., Donjacour, A., Cunha, G., & Baskin, L. (2004). Anatomical studies of the fibroblast growth factor-10 mutant, Sonic Hedge Hog mutant and androgen receptor mutant mouse genital tubercle. Advances in Experimental Medicine and Biology, 545, 123–148. https://doi.org/10.1007/978-1-4419-8995-6_8
Zheng, Z., Armfield, B., & Cohn, M. (2015). Timing of androgen receptor disruption and estrogen exposure underlies a spectrum of congenital penile anomalies. Proceedings of the National Academy of Sciences of the United States of America, 112(52), E7194-203. https://doi.org/10.1073/pnas.1515981112