
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 619
Title
Androgen receptor antagonism leads to delayed preputial separation via reduced fibroblast growth factor in genital-tubercle tissues
Short name
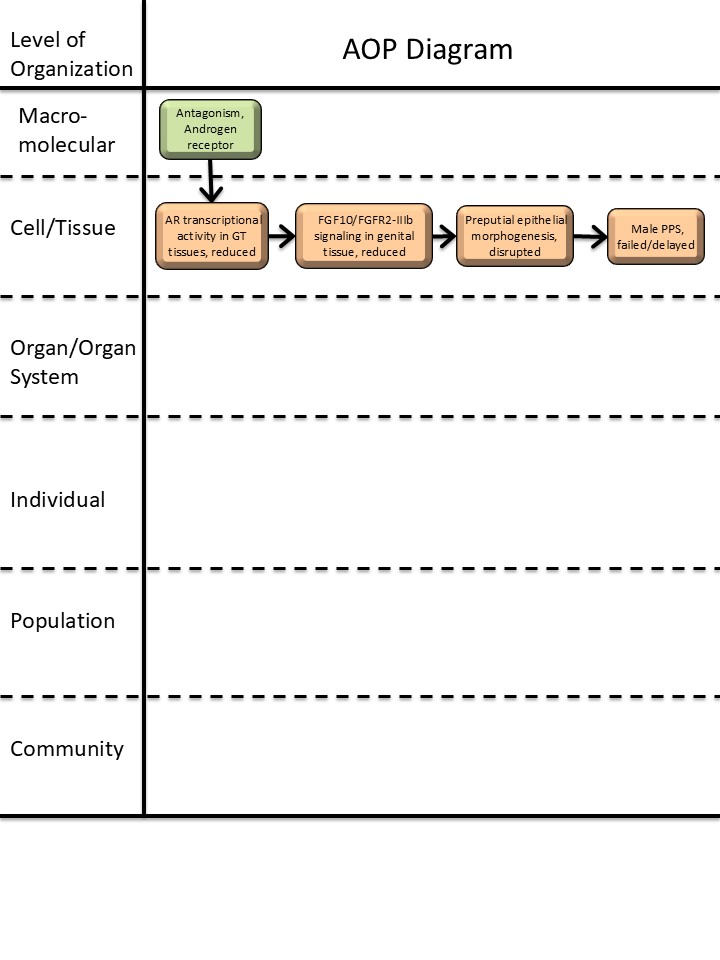
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 619, Androgen receptor antagonism leads to delayed preputial separation via reduced fibroblast growth factor in genital-tubercle tissues in tools offered by third parties.
Point of Contact
Contributors
- Travis Karschnik
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on July 08, 2026 15:53
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Antagonism, Androgen receptor | March 20, 2026 11:42 |
| Androgen receptor nuclear transcriptional activity in genital-tubercle tissues, reduced | December 18, 2025 11:35 |
| Fibroblast growth factor 10, fibroblast growth factor receptor 2 isoform IIIb signaling in genital tissue, reduced | December 18, 2025 11:43 |
| Preputial epithelial morphogenesis, disrupted | December 18, 2025 15:33 |
| Male preputial separation, failed/delayed | February 06, 2026 16:55 |
| Antagonism, Androgen receptor leads to AR transcriptional activity in GT tissues, reduced | April 08, 2026 13:13 |
| AR transcriptional activity in GT tissues, reduced leads to FGF10/FGFR2-IIIb signaling in genital tissue, reduced | May 07, 2026 15:54 |
| FGF10/FGFR2-IIIb signaling in genital tissue, reduced leads to Preputial epithelial morphogenesis, disrupted | June 04, 2026 15:18 |
| Preputial epithelial morphogenesis, disrupted leads to Male PPS, failed/delayed | July 07, 2026 17:16 |
| Dibutyl phthalate | November 29, 2016 18:42 |
| Flutamide | August 14, 2025 05:22 |
| Vinclozolin | May 14, 2020 11:28 |
| Procymidone | May 18, 2020 12:55 |
| Linuron | May 18, 2020 12:53 |
Abstract
This AOP describes how antagonism of the androgen receptor (AR) during genital tubercle (GT) development reduces AR-dependent nuclear transcriptional activity, thereby suppressing FGF10/FGFR2-IIIb signaling in genital tissues, disrupting preputial epithelial morphogenesis, and ultimately causing failed or delayed male preputial separation (PPS). The pathway is built on strong developmental genetics and toxicology evidence from rodent systems, particularly ex vivo mouse GT culture and in vivo rat anti-androgen studies. In mouse GT culture, flutamide caused dose-dependent downregulation of Fgfr2-IIIb and Fgf10, and DHT rescued the Fgfr2-IIIb and morphologic effects, indicating that the upstream AR event is causally linked to downstream FGF signaling in GT tissues (Petiot et al., 2005). Genetic evidence shows that Fgfr2-IIIb and Fgf10 are required for normal urethral epithelium maintenance and ventral prepuce development; loss of this signaling causes severe hypospadias-like malformations and failure of epithelial maturation (Petiot et al., 2005; Revest et al., 2001). Rodent developmental anti-androgen studies demonstrate that disruption of androgen signaling during the masculinization programming window delays PPS and induces persistent genital malformations, supporting the biological and regulatory relevance of the terminal adverse outcome (Mylchreest et al., 1999; Welsh et al., 2008; Sinclair et al., 2017). Gaps remain in the direct chromatin-level demonstration of AR regulation of Fgfr2 in GT tissue and in fully quantifying how specific morphogenetic lesions translate to PPS delay. Overall, the AOP has moderate-to-high confidence and is well suited for developmental hazard identification, prioritization, and mechanistic interpretation of anti-androgenic chemicals.
AOP Development Strategy
Context
This AOP was as part of an Environmental Protection Agency effort to develop AOPs that establish scientifically supported causal linkages between alternative endpoints measured using new approach methodologies (NAMs) and guideline apical endpoints measured in Tier 1 and Tier 2 test guidelines (U.S. EPA, 2024) employed by the Endocrine Disruptor Screening Program (EDSP). A series of key events that represent significant, measurable, milestones connecting molecular initiation to apical endpoints indicative of adversity were identified based on scientific review articles and empirical studies. Additionally, scientific evidence supporting the causal relationships between each pair of key events was assembled through a combination of expert knowledge and AI-assisted literature search and synthesis. Specifically, a combination of Claude (Anthropic) using Sonnet 4.6, and EPA AI, using GPT5, was used to identify, retrieve, and summarize relevant primary and secondary literature, and to draft the initial content of this AOP page. Users of this AOP are advised that AI-assisted evidence assembly may introduce selection bias or gaps in coverage that differ from a fully systematic human-conducted review, and independent verification of the evidence base is encouraged. A copy of the initial prompt is attached at the bottom of the Abstract section of this AOP.
Strategy
The scope of the aforementioned EPA project was to develop AOP(s) relevant to apical endpoints observed in the test guidelines, based on mechanisms consistent with empirical studies. The literature used to support this AOP and its constituent pages began with the test guidelines and followed to primary, secondary, and/or tertiary works concerning the relevant underlying biology. KE and KER page creation and re-use was determined using Handbook principles where page re-use was preferred.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 26 | Antagonism, Androgen receptor | Antagonism, Androgen receptor |
| KE | 2393 | Androgen receptor nuclear transcriptional activity in genital-tubercle tissues, reduced | AR transcriptional activity in GT tissues, reduced |
| KE | 2399 | Fibroblast growth factor 10, fibroblast growth factor receptor 2 isoform IIIb signaling in genital tissue, reduced | FGF10/FGFR2-IIIb signaling in genital tissue, reduced |
| KE | 2400 | Preputial epithelial morphogenesis, disrupted | Preputial epithelial morphogenesis, disrupted |
| AO | 2401 | Male preputial separation, failed/delayed | Male PPS, failed/delayed |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Embryo | High |
| Foetal | High |
| Fetal to Parturition | High |
| Development | High |
| Juvenile | High |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Overall Assessment of the AOP
Overall confidence in this AOP is moderate to high. Biological plausibility is high across all KERs, because the pathway follows well-established steroid receptor biology and canonical epithelial-mesenchymal FGF signaling during genital development (Gelmann, 2002; Itoh & Ornitz, 2011; Ohuchi et al., 2000; Quigley et al., 1995). Empirical support is also strong: the AR→FGF10/FGFR2-IIIb link is directly supported by flutamide/DHT rescue data in GT culture, the FGF signaling→morphogenesis link is strongly supported by loss-of-function mouse studies, and the final morphogenesis→PPS link is supported by rat developmental anti-androgen studies and histologic analyses of preputial fusion defects (Petiot et al., 2005; Revest et al., 2001; Gredler et al., 2015; Mylchreest et al., 1999; Sinclair et al., 2017).
The primary uncertainties are: (1) direct AR binding to the Fgfr2 regulatory region in GT tissue has not been experimentally confirmed; (2) Fgf10 regulation by AR may be indirect in GT tissue; and (3) some PPS studies do not measure preputial histology in the same animals, making the morphogenesis-to-PPS bridge partly inferential rather than directly observed (Petiot et al., 2005; Sinclair et al., 2017). These gaps do not undermine the overall pathway, but they do limit the precision of quantitative prediction.
Domain of Applicability
Taxonomic Applicability
The AOP is best supported in rodents, especially mouse and rat. Mouse ex vivo and genetic studies provide the strongest mechanistic support for the AR→FGF10/FGFR2-IIIb→preputial morphogenesis sequence, while rat developmental toxicology provides the clearest evidence for delayed PPS as the adverse outcome (Petiot et al., 2005; Harada et al., 2015; Gredler et al., 2015; Mylchreest et al., 1999; Sinclair et al., 2017). Human developmental and histologic studies support translational relevance but do not yet provide direct evidence for the full pathway (Kim et al., 2002; Beleza-Meireles et al., 2007; Carmichael et al., 2013; Haid et al., 2020).
Lifestage Applicability
The pathway is operative during embryonic and fetal external genital development, with the adverse outcome manifesting postnatally in juvenile males as delayed PPS. The critical androgen-sensitive window precedes or overlaps external genital morphogenesis and is distinct from the later postnatal timing of PPS scoring in rat toxicology studies (Mylchreest et al., 1999; Welsh et al., 2008; Seifert et al., 2008; Sinclair et al., 2017).
Sex Applicability
The molecular events can be detected in both sexes in organ culture, but the adverse outcome is male-specific because PPS is a male external genital endpoint. Accordingly, the AOP is most directly applicable to males, with upstream mechanistic relevance in mixed-sex developmental contexts (Petiot et al., 2005; Seifert et al., 2008).
Essentiality of the Key Events
|
Event |
Direct Evidence |
Indirect Evidence |
No experimental evidence |
Contradictory experimental evidence |
|
MIE: Antagonism, Androgen receptor |
*** |
** |
||
|
KE1: Androgen receptor nuclear transcriptional activity in genital-tubercle tissues, reduced |
*** |
* |
||
|
KE2: Fibroblast growth factor 10, fibroblast growth factor receptor 2 isoform IIIb signaling in genital tissue, reduced |
**** |
|||
|
KE3: Preputial epithelial morphogenesis, disrupted |
*** |
** |
||
|
AO: Male preputial separation, failed/delayed |
**** |
MIE: Antagonism, Androgen receptor
Direct evidence is strong because pharmacological AR antagonism with flutamide in GT organ culture suppresses downstream Fgfr2-IIIb/Fgf10 expression and can be rescued by DHT, demonstrating that antagonism of AR initiates the pathway (Petiot et al., 2005; Furutani et al., 2002). Indirect evidence also comes from developmental anti-androgen studies that reproduce the downstream phenotype in vivo (Mylchreest et al., 1999; Welsh et al., 2008; Sinclair et al., 2017).
KE1: Androgen receptor nuclear transcriptional activity in genital-tubercle tissues, reduced
Direct evidence is strong from flutamide-induced suppression of AR-dependent transcript expression in GT organ culture and DHT rescue of the response (Petiot et al., 2005). Indirect evidence includes in vivo anti-androgen studies that are consistent with reduced AR-driven developmental programs in the GT and downstream genital morphogenesis defects (Seifert et al., 2008; Mylchreest et al., 1999).
KE2: FGF10/FGFR2-IIIb signaling in genital tissue, reduced
Direct evidence is strong because Fgfr2-IIIb and Fgf10 are downregulated by flutamide in GT culture in a dose-dependent manner, and genetic loss of Fgfr2-IIIb or Fgf10 causes severe external genital defects and failure of urethral epithelial maturation (Petiot et al., 2005; Revest et al., 2001; Gredler et al., 2015).
KE3: Preputial epithelial morphogenesis, disrupted
Direct evidence is strong from mouse loss-of-function and tissue-specific FGF signaling studies showing abnormal urethral/preputial epithelial maturation and ventral prepuce closure defects when this pathway is impaired (Petiot et al., 2005; Gredler et al., 2015; Harada et al., 2015). Indirect evidence also comes from anti-androgen developmental studies in rats, which produce preputial/genital malformations consistent with disrupted morphogenesis before PPS is delayed (Yoshimura et al., 2004, 2005; Mylchreest et al., 1999).
AO: Male preputial separation, failed/delayed
Direct evidence is strong because developmental anti-androgen exposure delays PPS in rats and, in some cases, ventral defects prevent complete separation altogether (Mylchreest et al., 1999; Welsh et al., 2008; Sinclair et al., 2017; Yoshimura et al., 2004, 2005).
Evidence Assessment
KER 1: Antagonism, Androgen receptor → Androgen receptor nuclear transcriptional activity in genital-tubule tissues, reduced
Biological plausibility: High. AR is a ligand-activated nuclear steroid receptor, and antagonists block its transactivation function (Gelmann, 2002; Quigley et al., 1995).
Empirical support: High. Flutamide reduced downstream AR-responsive transcriptional output in GT culture, and DHT reversed the effect (Petiot et al., 2005).
Quantitative understanding: Moderate. The antagonist-dependent suppression is dose-responsive in GT tissue, but the quantitative relationship is not fully parameterized for in vivo extrapolation (Petiot et al., 2005).
KER 2: Androgen receptor nuclear transcriptional activity in genital-tubule tissues, reduced → Fibroblast growth factor 10, fibroblast growth factor receptor 2 isoform IIIb signaling in genital tissue, reduced
Biological plausibility: High. AR-dependent transcription can regulate developmental growth factor signaling, and FGF10/FGFR2-IIIb is a canonical epithelial-mesenchymal signaling pair in genital development (Itoh & Ornitz, 2011; Ohuchi et al., 2000; Petiot et al., 2005).
Empirical support: High. Flutamide caused dose-dependent reduction of Fgfr2-IIIb and Fgf10, and DHT rescued Fgfr2-IIIb expression and normal morphology; an in silico ARE was identified in the Fgfr2 promoter, though direct AR chromatin binding was not shown (Petiot et al., 2005).
Quantitative understanding: Moderate. Dose-response relationships were demonstrated in organ culture, but the quantitative mechanistic threshold between AR activity and FGF pathway output is not fully resolved (Petiot et al., 2005).
KER 3: Fibroblast growth factor 10, fibroblast growth factor receptor 2 isoform IIIb signaling in genital tissue, reduced → Preputial epithelial morphogenesis, disrupted
Biological plausibility: High. FGF10/FGFR2-IIIb controls epithelial proliferation, differentiation, and morphogenesis in multiple organs, including the GT (Harada et al., 2015; Itoh & Ornitz, 2011; Ohuchi et al., 2000).
Empirical support: High. Fgfr2-IIIb-null mice exhibit severe urethral developmental defects, premature arrest of epithelial proliferation, and failed preputial morphogenesis; similar phenotypes occur when Fgf10 signaling is disrupted (Petiot et al., 2005; Revest et al., 2001; Gredler et al., 2015).
Quantitative understanding: Moderate. The genotype-phenotype relationship is strong, but a formal quantitative model linking signaling intensity to morphogenetic severity is not available.
KER 4: Preputial epithelial morphogenesis, disrupted → Male preputial separation, failed/delayed
Biological plausibility: High. PPS requires normal preputial epithelial fusion/canalization and subsequent separation; therefore, disruption of morphogenesis is a direct precursor to delayed or failed PPS (Gairdner, 1949; Sinclair et al., 2017; Yoshimura et al., 2004, 2005).
Empirical support: High. Developmental anti-androgen exposure and morphologic disruption of the preputial region are consistently associated with delayed or failed PPS in rats (Mylchreest et al., 1999; Welsh et al., 2008; Sinclair et al., 2017).
Quantitative understanding: Moderate. PPS timing is readily measured, but the exact quantitative mapping from specific epithelial lesions to day-of-separation shifts remains incompletely defined.
Known Modulating Factors
|
Modulating Factor (MF) |
Influence or Outcome |
KER(s) involved |
Quantitative Understanding |
|
Developmental timing of exposure |
Exposure during the masculinization programming window has the greatest impact on genital morphogenesis and PPS |
3705, 3706, 3707, 3708 |
Moderate |
|
Dose of AR antagonist |
Higher antagonist doses produce greater suppression of AR-dependent transcription and downstream FGF signaling |
3705, 3706 |
Moderate |
|
Sex / androgen milieu |
Male fetuses/juveniles are more susceptible to the adverse outcome because PPS is androgen-dependent |
3705, 3706, 3707, 3708 |
Moderate |
|
Genetic background / strain |
Severity of genital malformations and PPS delay varies among rodent strains |
3707, 3708 |
Low to moderate |
|
General growth delay / body weight |
Non-specific developmental delay can shift PPS timing and confound interpretation |
3708 |
Moderate |
The main modulating factors are developmental timing, sex-specific androgen milieu, antagonist dose, genetic background, and general growth delay. These factors are most relevant to the AR→FGF signaling and morphogenesis→PPS segments of the pathway, where they influence the magnitude and timing of the downstream phenotype (Mylchreest et al., 1999; Welsh et al., 2008; Sinclair et al., 2017).
Quantitative Understanding
Quantitative understanding is moderate for the early KERs and the AO, and lower for the integrated pathway as a whole. There is clear dose-dependent suppression of Fgfr2-IIIb and Fgf10 by flutamide in GT culture and a clear phenotypic association between developmental anti-androgen exposure and delayed PPS in rats, but a fully parameterized quantitative model linking AR antagonism to PPS timing is not yet available (Petiot et al., 2005; Mylchreest et al., 1999; Welsh et al., 2008). The pathway is therefore more suitable for screening and prioritization than for stand-alone quantitative risk prediction.
Considerations for Potential Applications of the AOP (optional)
This AOP is well suited for regulatory screening, hazard prioritization, and weight-of-evidence evaluation of anti-androgenic chemicals. It can support grouping and read-across for substances with AR antagonism, assist in interpreting developmental toxicity studies, and help connect in vitro AR assays to in vivo male reproductive developmental outcomes. The pathway is also useful for identifying data gaps when a chemical perturbs AR signaling but has not yet been tested for GT or PPS endpoints. Application to human risk assessment should remain cautious because the strongest evidence derives from rodent systems and because human external genital development differs in timing and anatomy (Kim et al., 2002; Beleza-Meireles et al., 2007; Carmichael et al., 2013; Sharpe, 2020).
References
Beleza-Meireles, A., Lundberg, F., Lagerstedt, K., & others. (2007). FGFR2, FGF8, FGF10 and BMP7 as candidate genes for hypospadias. European Journal of Human Genetics, 15(4), 405–410. https://doi.org/10.1038/sj.ejhg.5201777
Carmichael, S. L., Ma, C., Choudhry, S., Lammer, E. J., Witte, J. S., & Shaw, G. M. (2013). Hypospadias and genes related to genital tubercle and early urethral development. Journal of Urology, 190(5), 1884–1892. https://doi.org/10.1016/j.juro.2013.05.061
Furutani, T., Watanabe, T., Tanimoto, K., Hashimoto, T., Koutoku, H., Kudoh, M., Shimizu, Y., Kato, S., & Shikama, H. (2002). Stabilization of androgen receptor protein is induced by agonist, not by antagonists. Biochemical and Biophysical Research Communications, 294, 779–784.
Gelmann, E. P. (2002). Molecular biology of the androgen receptor. Journal of Clinical Oncology, 20(13), 3001–3015. https://doi.org/10.1200/JCO.2002.10.018
Gredler, M. L., Seifert, A. W., & Cohn, M. J. (2015). Tissue-specific roles of Fgfr2 in development of the external genitalia. Development, 142(12), 2203–2212. https://doi.org/10.1242/dev.119891
Haid, B., Pechriggl, E., Nägele, F., et al. (2020). FGF8, FGF10 and FGF receptor 2 in foreskin of children with hypospadias: An analysis of immunohistochemical expression patterns and gene transcription. Journal of Pediatric Urology, 16(1), 41.e1–41.e10. https://doi.org/10.1016/j.jpurol.2019.10.007
Harada, M., Omori, A., Nakahara, C., Nakagata, N., Akita, K., & Yamada, G. (2015). Tissue-specific roles of FGF signaling in external genitalia development. Developmental Dynamics, 244(6), 759–773. https://doi.org/10.1002/dvdy.24277
Itoh, N., & Ornitz, D. M. (2011). Fibroblast growth factors: From molecular evolution to roles in development, metabolism and disease. Journal of Biochemistry, 149(2), 121–130.
Kim, K. S., Liu, W., Cunha, G. R., Russell, D. W., Huang, H., Shapiro, E., & Baskin, L. S. (2002). Expression of the androgen receptor and 5α-reductase type 2 in the developing human fetal penis and urethra. Cell and Tissue Research, 307(2), 145–153.
Mylchreest, E., Sar, M., Wallace, D. G., & Foster, P. M. D. (1999). Fetal testosterone insufficiency causes reproductive tract abnormalities in male rats. Toxicological Sciences, 52(1), 116–127.
Ohuchi, H., Hori, Y., Yamasaki, M., Harada, H., Sekine, K., Kato, S., & Itoh, N. (2000). FGF10 acts as a major ligand for FGF receptor 2 IIIb in mouse multi-organ development. Biochemical and Biophysical Research Communications, 277(3), 643–649. https://doi.org/10.1006/bbrc.2000.3721
Petiot, A., Perriton, C. L., Dickson, C., & Cohn, M. J. (2005). Development of the mammalian urethra is controlled by Fgfr2-IIIb. Development, 132(10), 2441–2450. https://doi.org/10.1242/dev.01778
Quigley, C. A., De Bellis, A., Marschke, K. B., el-Awady, M. K., Wilson, E. M., & French, F. S. (1995). Androgen receptor defects: Historical, clinical, and molecular perspectives. Endocrine Reviews, 16(3), 271–321. https://doi.org/10.1210/edrv-16-3-271
Revest, J. M., Spencer-Dene, B., Kerr, K., De Moerlooze, L., Rosewell, I., & Dickson, C. (2001). Fibroblast growth factor receptor 2-IIIb acts upstream of Shh and Fgf4 and is required for limb bud maintenance but not for the induction of Fgf8, Fgf10, Msx1, or Bmp4. Developmental Biology, 231(1), 47–62. https://doi.org/10.1006/dbio.2000.0144
Seifert, A. W., Harfe, B. D., & Cohn, M. J. (2008). Cell lineage analysis demonstrates an endodermal origin of the distal urethra and perineum. Developmental Biology, 318(1), 143–152. https://doi.org/10.1016/j.ydbio.2008.03.017
Sharpe, R. M. (2020). Androgens and the masculinization programming window: Human-rodent differences. Biochemical Society Transactions, 48(4), 1725–1735. https://doi.org/10.1042/BST20200200
Sinclair, A. W., Cao, M., Pask, A., Baskin, L. S., & Cunha, G. R. (2017). Flutamide-induced hypospadias in rats: A critical assessment. Differentiation, 94, 37–57. https://doi.org/10.1016/j.diff.2016.12.001
Welsh, M., Saunders, P. T. K., Fisken, M., Scott, H. M., Hutchison, G. R., Smith, L. B., & Sharpe, R. M. (2008). Identification in rats of a programming window for reproductive tract masculinization, disruption of which leads to hypospadias and cryptorchidism. The Journal of Clinical Investigation, 118(4), 1479–1490. https://doi.org/10.1172/JCI34241