
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 331
Title
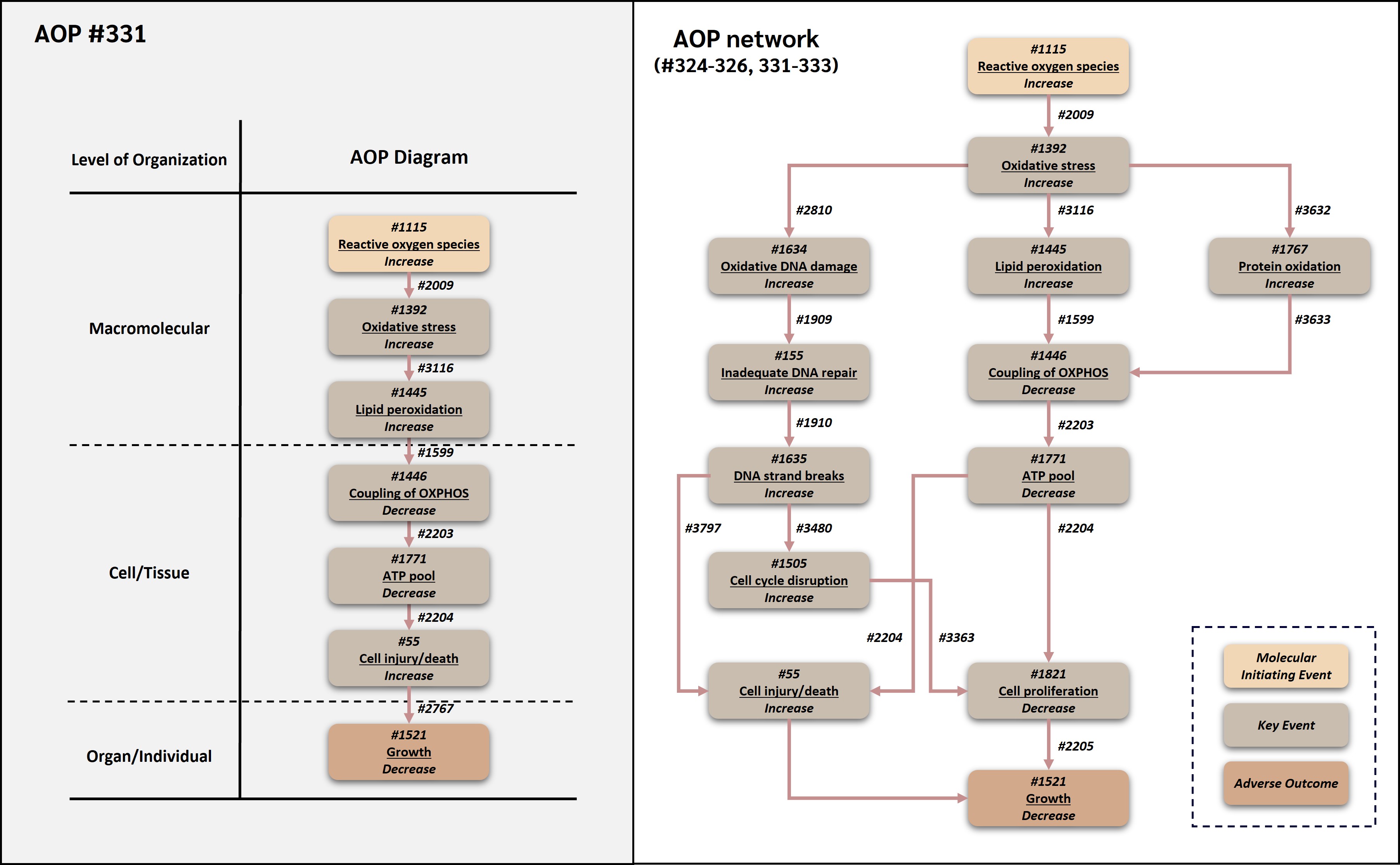
Reactive oxygen species leading to growth inhibition via lipid peroxidation and cell death
Short name
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 331, Reactive oxygen species leading to growth inhibition via lipid peroxidation and cell death in tools offered by third parties.
Point of Contact
Contributors
- You Song
Coaches
- Shihori Tanabe
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on June 20, 2026 05:38
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Decrease, Growth | July 06, 2022 07:36 |
| Increase, Reactive oxygen species | June 12, 2025 01:27 |
| Increase, Oxidative Stress | February 11, 2026 07:05 |
| Increase, Lipid peroxidation | June 23, 2026 06:46 |
| Decrease, Coupling of oxidative phosphorylation | November 07, 2025 05:15 |
| Decrease, Adenosine triphosphate pool | June 14, 2021 13:40 |
| Increase, Cell injury/death | May 27, 2024 07:23 |
| Increase, ROS leads to Increase, Oxidative Stress | June 23, 2026 06:57 |
| Increase, Oxidative Stress leads to Increase, LPO | June 23, 2026 06:59 |
| Increase, LPO leads to Decrease, Coupling of OXPHOS | June 23, 2026 07:21 |
| Decrease, Coupling of OXPHOS leads to Decrease, ATP pool | July 06, 2022 07:39 |
| Decrease, ATP pool leads to Cell injury/death | June 23, 2026 07:40 |
| Cell injury/death leads to Decrease, Growth | June 23, 2026 07:54 |
| Ultraviolet B radiation | April 15, 2017 16:04 |
| Hydrogen peroxide | May 19, 2019 17:21 |
| Paraquat | November 29, 2016 18:42 |
| tert-Butyl hydroperoxide | May 19, 2019 17:24 |
| Heavy metals (cadmium, lead, copper, iron, nickel) | October 25, 2021 03:21 |
| Silver | February 03, 2022 11:20 |
| Silver nanoparticles | February 15, 2017 03:19 |
| Ionizing Radiation | May 07, 2019 12:12 |
Abstract
This adverse outcome pathway (AOP 331) describes a linear route by which increased reactive oxygen species (ROS) can lead to decreased organismal growth through lipid peroxidation-mediated mitochondrial bioenergetic impairment and increased cell injury/death. In this AOP, increased ROS is treated operationally as the molecular initiating event because it represents the earliest common measurable redox perturbation shared by many chemical and non-chemical stressors within the broader ROS-growth AOP network. Increased ROS leads to oxidative stress, which promotes lipid peroxidation. Oxidative damage to membrane lipids can impair mitochondrial membrane integrity and coupling of oxidative phosphorylation (OXPHOS). Decreased OXPHOS coupling reduces ATP production, and insufficient ATP availability can compromise membrane homeostasis, ion transport, biosynthesis, stress-response capacity, and execution of regulated cell death pathways, ultimately resulting in increased cell injury/death. Increased loss of viable cells, particularly in developing, growing, or regenerating tissues and organisms, can contribute to decreased growth.
AOP 331 reuses and connects established AOP-Wiki components from several AOP contexts. The upstream ROS and oxidative stress segment is associated with AOP 478, in which deposition of energy leads to oxidative stress through increased free radical generation (AOP-Wiki, 2026a). The lipid peroxidation and mitochondrial bioenergetic segment is connected to the oxidative stress and mitochondrial impairment logic represented in the broader ROS-growth AOP network, while the KER from decreased coupling of OXPHOS to decreased ATP pool is directly associated with AOP 263, an OECD-published AOP that causally links uncoupling of OXPHOS to growth inhibition through ATP depletion and decreased cell proliferation (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). AOP 331 differs from AOP 326 by routing ATP depletion through increased cell injury/death rather than decreased cell proliferation. This terminal cellular injury module is supported by reuse of the broadly shared AOP-Wiki KE 'Increase, Cell injury/death' and by its occurrence in several AOPs, including AOPs 12, 13, 17, 38, and 48, where cell injury/death is used as an intermediate or downstream KE in neurotoxicity, oxidative stress, fibrosis, and excitotoxicity contexts (AOP-Wiki, 2026c-g). The AOP is relevant to environmental and human health contexts because ROS production, lipid peroxidation, mitochondrial ATP production, cell viability, and growth are conserved biological processes. It can support mechanistic interpretation of oxidative stress-mediated growth impairment, assay selection, chemical prioritization, integrated approaches to testing and assessment (IATA), and quantitative AOP development for oxidative and mitochondrial toxicity.
Acknowledgement
This project was funded by the Research Council of Norway (RCN), grant no. RCN-315929 “EXPECT: In silico and experimental screening platform for characterizing environmental impact of industry development in the Arctic” (https://www.niva.no/en/projects/expect), the European Partnership for the Assessment of Risks from Chemicals (PARC) through European Union’s Horizon Europe research and innovation programme (Grant Agreement No 101057014, and supported by the NIVA Computational Toxicology Program, NCTP (https://www.niva.no/en/featured-pages/nctp, grant. No. RCN-342628).
AI disclosure
Artificial intelligence (AI) tools were used to support literature prioritization, review and AOP-Wiki page preparation in this work. AOP-helpFinder was used for automated literature mining, and ChatGPT (OpenAI) was used as an auxiliary tool for title and abstract screening, extraction of study metadata, and identification of potential weight-of-evidence indicators. AI-assisted outputs were used only to organize and prioritize information and were verified against the original sources by the authors before inclusion. Additional AI assistance was used for formatting, copy-editing, citation cross-checking, and harmonization of the AOP-Wiki pages. All scientific interpretations, weight-of-evidence judgments, final wording, and conclusions were determined and approved by the authors, who take full responsibility for the content and integrity of the work.
AOP Development Strategy
Context
ROS are continuously formed during aerobic metabolism and can also be generated in response to environmental stressors. At controlled levels, ROS participate in redox signaling, whereas excessive ROS can disturb redox homeostasis and initiate oxidative stress (Schieber and Chandel, 2014; Sies et al., 2017). Lipid membranes are important targets of oxidative attack because phospholipids containing polyunsaturated fatty acids can undergo radical-driven peroxidation. Lipid peroxidation generates lipid hydroperoxides and secondary reactive aldehydes, including malondialdehyde and 4-hydroxy-2-nonenal, which can propagate oxidative injury and alter membrane-associated protein and organelle function (Ayala et al., 2014).
AOP 331 was developed to represent the lipid peroxidation and cell injury/death-driven linear route within the broader ROS-growth AOP network. This route was selected because lipid peroxidation is a well-established consequence of oxidative stress and because mitochondrial membranes are central determinants of OXPHOS coupling. Peroxidative modification of mitochondrial membrane lipids can alter membrane fluidity, proton leak, respiratory control, and mitochondrial membrane potential, providing a mechanistically coherent bridge from oxidative stress to impaired ATP production (Murphy, 2009; Nicholls and Ferguson, 2013; Ouillon et al., 2021). ATP depletion is a well-established contributor to loss of cell viability because cellular survival depends on ATP-dependent ion gradients, membrane repair, protein turnover, stress-response pathways, and the execution of regulated death processes. Depletion of ATP can shift cells from adaptive responses to injury and death, and severe ATP loss can affect the mode of cell death (Leist et al., 1997; Bonora et al., 2012).
The AOP was also developed to take advantage of existing AOP-Wiki modularity. The upstream oxidative stress context is associated with AOP 478, while the OXPHOS-to-ATP KER is associated with AOP 263 (AOP-Wiki, 2026a,b; OECD, 2022). The cell injury/death KE is a highly reusable AOP-Wiki KE and appears across several established AOPs. AOP 17 explicitly includes oxidative stress leading to cell injury/death and also includes several KERs involving cell injury/death and neuroinflammation (AOP-Wiki, 2026e). AOP 48 includes mitochondrial dysfunction leading to cell injury/death in an excitotoxicity context (AOP-Wiki, 2026g). AOP 38 uses cell injury and cell death as key early tissue-level consequences of protein alkylation leading to fibrosis (AOP-Wiki, 2026f). AOPs 12 and 13 also use cell injury/death in neurodegeneration and synaptogenesis-related contexts (AOP-Wiki, 2026c,d). These associations support the reuse of Event 55 as a generic, modular cellular KE downstream of multiple upstream stressors and upstream of multiple adverse outcomes.
Strategy
AOP 331 was developed using the principles described in OECD AOP guidance, including modular description of KEs and KERs, reuse of existing AOP-Wiki content where appropriate, evidence evaluation using biological plausibility, empirical support, essentiality, and quantitative understanding, and clear description of the biological domain of applicability (OECD, 2018, 2021). The aim was to assemble a focused linear pathway from reusable AOP-Wiki elements rather than to create an isolated de novo pathway. This is important because AOP 331 is one branch of the broader ROS-growth AOP network and because its KEs overlap with oxidative stress, mitochondrial dysfunction, cellular energy metabolism, cell injury/death, and growth-related AOPs.
Reuse of existing AOP-Wiki content was considered at the outset. AOP 478 was reviewed because it provides an AOP-Wiki precedent for oxidative stress as a central KE downstream of free radical generation and energy deposition. AOP 263 was reviewed because it provides an OECD-published downstream bioenergetics module in which decreased coupling of OXPHOS leads to decreased ATP pool and subsequently growth inhibition, although in AOP 263 the terminal cellular route proceeds through decreased cell proliferation rather than cell injury/death. AOPs 12, 13, 17, 38, and 48 were reviewed because they demonstrate repeated reuse of Event 55, 'Increase, Cell injury/death', across different biological contexts and provide support for treating cell injury/death as a modular KE that can connect distinct upstream mechanisms to downstream tissue or organism-level outcomes. AOP 296 was reviewed during development of the broader ROS-growth network to ensure that oxidative stress and macromolecular damage modules were harmonized with existing oxidative damage content, although AOP 331 specifically follows the lipid peroxidation and bioenergetic injury branch rather than the oxidative DNA damage branch.
The evidence base was assembled through an AI-human hybrid workflow. First, search terms were developed for each KE, including KE names, synonyms, endpoint names, assay terms, taxa, and representative stressors. AOP-helpFinder was used to search PubMed for co-occurrence between key events and related biological concepts, and the exported outputs included PMIDs, titles, abstracts, and matched KE terms (Carvaillo et al., 2019; Jornod et al., 2022). The exported records were subjected to overlap analysis to remove redundant hits and to filter taxa-related or clearly irrelevant literature.
Second, ChatGPT (OpenAI, San Francisco, CA, USA)-assisted screening was used as an auxiliary prioritization step. The LLM was used to pre-screen titles and abstracts, extract study metadata including stressor, species, biological system, dose or concentration, and exposure time, identify evidence types such as biological plausibility, empirical support, and essentiality, and flag weight-of-evidence indicators such as dose-response concordance, temporal concordance, incidence concordance, and intervention evidence. The LLM output was used to classify studies as high relevance, medium relevance, or low/not relevant. High-relevance studies were retrieved for full-text review, medium-relevance studies were reserved as supporting evidence, and low-relevance studies were documented as low priority or excluded.
Third, full-text review and expert curation were used to verify all evidence before inclusion in the AOP. LLM-assisted full-text review was used only to organize candidate evidence; all extracted information was checked manually against the original text. Expert review was then used to populate KER evidence tables with methods, endpoints, results, weight-of-evidence category, and references. Final weight-of-evidence evaluation was performed by expert judgment using biological plausibility, empirical support, essentiality, quantitative understanding, and identification of evidence gaps. Thus, the development process combined text-mining and AI-assisted evidence handling with human expert verification and final decision-making.
In parallel with this workflow, targeted searches were conducted to fill specific evidence gaps for ROS, oxidative stress, lipid peroxidation, mitochondrial membrane potential, OXPHOS coupling, ATP depletion, cytotoxicity, cell death, and growth inhibition. Studies were prioritized when they measured two or more KEs in the same biological system, reported dose or concentration and exposure time, or provided evidence relevant to dose-response, temporal, or incidence concordance. Mechanistic reviews and OECD reports were used primarily to support biological plausibility, while primary experimental studies were used to support empirical concordance wherever possible.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 1115 | Increase, Reactive oxygen species | Increase, ROS |
| KE | 1392 | Increase, Oxidative Stress | Increase, Oxidative Stress |
| KE | 1445 | Increase, Lipid peroxidation | Increase, LPO |
| KE | 1446 | Decrease, Coupling of oxidative phosphorylation | Decrease, Coupling of OXPHOS |
| KE | 1771 | Decrease, Adenosine triphosphate pool | Decrease, ATP pool |
| KE | 55 | Increase, Cell injury/death | Cell injury/death |
| AO | 1521 | Decrease, Growth | Decrease, Growth |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Increase, ROS leads to Increase, Oxidative Stress | adjacent | High | Moderate |
| Increase, Oxidative Stress leads to Increase, LPO | adjacent | High | Moderate |
| Increase, LPO leads to Decrease, Coupling of OXPHOS | adjacent | High | Moderate |
| Decrease, Coupling of OXPHOS leads to Decrease, ATP pool | adjacent | High | High |
| Decrease, ATP pool leads to Cell injury/death | adjacent | High | Moderate |
| Cell injury/death leads to Decrease, Growth | adjacent | High | Moderate |
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| All life stages | Moderate |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Unspecific | Moderate |
Overall Assessment of the AOP
The overall weight of evidence supporting AOP 331 is considered moderate. Biological plausibility is high for all six KERs in the pathway. The upstream oxidative stress, lipid peroxidation, and OXPHOS uncoupling sequence follows a well-established mechanistic logic, and the connection from ATP depletion to cell injury/death is supported by the fundamental dependence of cellular survival on adequate energy supply. The cell injury/death-to-growth relationship is reinforced by the broad reuse of Event 55 (Increase, Cell injury/death) as a modular KE across endorsed AOPs 12, 13, 17, 38, and 48 (AOP-Wiki, 2026a-e). The OXPHOS-to-ATP module is directly associated with OECD-endorsed AOP 263 and contributes high biological plausibility and strong quantitative understanding for this segment (OECD, 2022; Song and Villeneuve, 2021). Empirical support is high for the ROS-to-oxidative-stress and oxidative-stress-to-lipid-peroxidation relationships, moderate for the lipid-peroxidation-to-OXPHOS link, and moderate to high for the ATP-depletion-to-cell-death and OXPHOS-to-ATP relationships. The cell death-to-growth relationship has moderate empirical support, as direct concurrent measurement of cell injury/death and organismal growth is less common across the available literature. Essentiality is rated moderate to high overall, with the strongest direct evidence for the AOP 263 bioenergetics segment. Quantitative understanding is highest for the OXPHOS-to-ATP KER and low to moderate elsewhere. The main uncertainties are the quantitative thresholds governing the lipid-peroxidation-to-OXPHOS transition, the severity-dependent mode of cell death triggered by ATP depletion, and the extent to which cell injury/death versus reduced proliferation drives growth impairment in specific biological contexts. AOP 331 is most appropriate for mechanistic interpretation of cytotoxic growth impairment caused by oxidative lipid damage, IATA development, and chemical prioritisation (OECD, 2018; Becker et al., 2015).
Domain of Applicability
The domain of applicability for AOP 331 is broad across aerobic eukaryotic organisms in which ROS generation, oxidative stress responses, lipid peroxidation, mitochondrial oxidative phosphorylation, ATP-dependent homeostasis, cell injury/death, and growth are biologically relevant. The AOP is most applicable to taxa and life stages in which growth depends strongly on maintenance of viable cell number, tissue condition, and mitochondrial energy supply. This includes algae, aquatic invertebrates, fish embryos and juveniles, mollusks, and mammalian or human cell systems.
The stressor domain includes direct ROS generators, redox-cycling chemicals, metals, nanoparticles, mitochondrial toxicants, hypoxia-reoxygenation, and radiation. Because the MIE is defined operationally as increased ROS rather than as a chemical-specific molecular interaction, AOP 331 should be applied to stressors for which evidence supports increased ROS or oxidative stress and downstream concordance with lipid peroxidation, mitochondrial impairment, ATP depletion, cell injury/death, and decreased growth. Environmental factors such as oxygen availability, temperature, lipid composition, diet, nutrient status, and antioxidant capacity may modulate the pathway.
Essentiality of the Key Events
Essentiality is evaluated for the overall AOP based on whether preventing or modifying upstream KEs changes downstream KEs or the AO. Direct essentiality evidence is strongest for the OXPHOS to ATP relationship and for ATP dependence of cell viability. Essentiality for lipid peroxidation is biologically plausible and supported by intervention and association studies, but direct experiments showing that blocking lipid peroxidation prevents all downstream events are less common.
|
Key event |
Essentiality |
Rationale |
Experimental manipulation evidence (KE knock-out / inhibition / rescue) |
Uncertainties |
|
Event 1115: Reactive oxygen species, increased |
Moderate |
ROS scavenging and antioxidant interventions frequently attenuate oxidative stress and downstream lipid peroxidation in oxidative stress models (Schieber and Chandel, 2014; Sies et al., 2017). |
Indirect (stop/attenuation): antioxidant and ROS-scavenger pre-treatment reduces oxidative stress and downstream damage across oxidative-stress models (Schieber and Chandel, 2014; Sies et al., 2017). No selective single-source ROS knock-out is available. |
ROS also participate in normal signaling; increased ROS does not always progress to adversity if compensation occurs. |
|
Event 1392: Oxidative stress, increased |
Moderate to high |
Oxidative stress is required for lipid peroxidation when oxidant production exceeds antioxidant buffering. AOP 478 and AOP 17 support oxidative stress as a central KE downstream of free radical generation or decreased protection against oxidative stress (AOP-Wiki, 2026a,e). |
Indirect: modulation of antioxidant capacity alters progression to oxidative macromolecular damage; oxidative stress is the curated hub KE in endorsed AOP 478 (AOP-Wiki, 2026a; Carrothers et al., 2025). |
Oxidative stress is measured using several indirect biomarkers that may not be equivalent across systems. |
|
Event 1445: Lipid peroxidation, increased |
Moderate |
Lipid peroxidation can alter membrane properties and generate reactive aldehydes that affect mitochondrial function (Ayala et al., 2014). Dietary PUFA studies in Daphnia show higher lipid peroxidation with lower mitochondrial membrane potential (Moore et al., 2023). |
Indirect: antioxidant intervention attenuates lipid peroxidation in oxidative-stress models; direct block-and-rescue isolating LPO from other oxidative damage is uncommon (Murphy, 2009; Ouillon et al., 2021). |
Direct blocking experiments are limited; lipid peroxidation may be both a cause and consequence of mitochondrial dysfunction. |
|
Event 1446: Coupling of OXPHOS, decreased |
High |
The KER from decreased OXPHOS coupling to ATP depletion is associated with AOP 263, where restoration or removal of uncoupling supports a causal role for impaired coupling in ATP depletion (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). |
Direct (rescue): removal of uncouplers or restoration of coupling recovers mitochondrial membrane potential and ATP in the endorsed AOP 263 module (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). |
Mild uncoupling can sometimes reduce ROS generation and may be adaptive; severity and duration determine adversity. |
|
Event 1771: ATP pool, decreased |
High |
ATP is required for maintenance of ion gradients, membrane repair, cellular stress responses, and execution of regulated cell death pathways. Severe ATP depletion is a well-established determinant of cell injury/death mode and severity (Leist et al., 1997; Bonora et al., 2012). |
Indirect: ATP-restoration experiments reduce downstream injury/proliferation deficits; central KE in endorsed AOP 263 (Leist et al., 1997; Nicotera et al., 1998; OECD, 2022). |
Compensatory glycolysis can buffer ATP depletion; total ATP may reflect changing cell number in some assays. |
|
Event 55: Cell injury/death, increased |
Moderate |
Cell injury/death is a shared AOP-Wiki KE used in AOPs 12, 13, 17, 38, and 48. Loss of viable cells provides a plausible and broadly supported mechanism for reduced tissue or organismal growth (AOP-Wiki, 2026c-g). |
Indirect: ATP restoration/maintenance reduces injury in some systems, indicating energy-status dependence (Leist et al., 1997; Nicotera et al., 1998); widely reused modular KE (AOPs 12, 13, 17, 38, 48). |
Growth can also decrease through reduced proliferation, altered cell size, endocrine disruption, or energy allocation without overt cell death. |
|
Event 1521: Growth, decreased (AO) |
Not applicable (AO) |
Growth is the adverse outcome and is regulatory relevant across algae, aquatic invertebrate, fish, amphibian, and plant test systems. AOP 263 provides precedent for using decreased growth as an AO in a mitochondrial bioenergetics AOP (OECD, 2022; Song and Villeneuve, 2021). |
As the adverse outcome, essentiality is assessed for upstream KEs; AOP 263 provides precedent for decreased growth as an AO downstream of these modules (OECD, 2022; Song and Villeneuve, 2021). |
Growth is integrative and can arise through multiple interacting mechanisms. |
Evidence Assessment
Evidence assessment is organized by KER. Calls follow OECD weight-of-evidence considerations for biological plausibility, empirical support, and quantitative understanding (OECD, 2018, 2021).
Biological plausibility of KERs
|
KER |
Biological plausibility call |
Rationale |
|
Relationship 2009: ROS increase leads to oxidative stress increase |
High |
Oxidative stress reflects an imbalance between oxidant production and antioxidant capacity, and ROS are primary oxidant species in cellular redox biology (Schieber and Chandel, 2014; Sies et al., 2017). AOP 478 supports oxidative stress downstream of free radical generation (AOP-Wiki, 2026a). |
|
Relationship 3116: oxidative stress increase leads to lipid peroxidation increase |
High |
ROS can initiate peroxidation of polyunsaturated fatty acids in membranes, generating lipid hydroperoxides and reactive aldehydes such as MDA and 4-HNE (Ayala et al., 2014; Sies et al., 2017). |
|
Relationship 1599: lipid peroxidation increase leads to decreased coupling of OXPHOS |
High |
Mitochondrial coupling depends on inner mitochondrial membrane integrity. Lipid peroxidation can disrupt membrane properties, promote proton leak, alter membrane potential, and impair respiratory control (Murphy, 2009; Nicholls and Ferguson, 2013; Ouillon et al., 2021). |
|
Relationship 2203: decreased coupling of OXPHOS leads to decreased ATP pool |
High |
This relationship is associated with AOP 263. OXPHOS coupling is a major determinant of ATP production in aerobic eukaryotic cells; reduced coupling lowers ATP synthesis efficiency (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). |
|
Relationship 2768: decreased ATP pool leads to increased cell injury/death |
High |
ATP is required for survival, ion homeostasis, membrane repair, and regulated death processes. Severe ATP depletion can switch cellular outcomes toward necrosis or irreversible injury, while less severe depletion may permit apoptosis (Leist et al., 1997; Bonora et al., 2012). |
|
Relationship 2767: increased cell injury/death leads to decreased growth |
High |
Growth depends on viable cell number, tissue integrity, and biomass accumulation. Increased cell death or injury reduces the cellular basis for growth and can impair tissue or organismal development (Conlon and Raff, 1999). Cell injury/death is reused across AOPs 12, 13, 17, 38, and 48 (AOP-Wiki, 2026c-g). |
Empirical support for KERs
|
KER |
Empirical support call |
Rationale |
Inconsistencies or evidence gaps |
|
Relationship 2009: ROS increase leads to oxidative stress increase |
High |
Paraquat increased ROS and antioxidant enzyme responses in Chlorella vulgaris (Qian et al., 2009), and paraquat induced oxidative stress responses in Daphnia magna (Barata et al., 2005). AOP 478 reports extensive evidence linking free radical generation/energy deposition to oxidative stress (AOP-Wiki, 2026a). |
ROS is often transient and measured indirectly; oxidative stress biomarkers vary across assays and taxa. |
|
Relationship 3116: oxidative stress increase leads to lipid peroxidation increase |
High |
Copper increased antioxidant enzyme activity and MDA/TBARS in freshwater green microalgae (Knauert and Knauer, 2008). Paraquat induced lipid peroxidation in algae and Daphnia (Barata et al., 2005; Esperanza et al., 2015; Qian et al., 2009). Gamma radiation in Lemna minor induced a sequential oxidative stress to lipid peroxidation response upstream of mitochondrial membrane potential loss and cell death (Xie et al., 2019; Xie et al., 2022). |
MDA/TBARS endpoints can lack specificity; lipid peroxidation and antioxidant responses may have different time courses. |
|
Relationship 1599: lipid peroxidation increase leads to decreased coupling of OXPHOS |
Moderate |
Dietary PUFA manipulation in Daphnia showed higher lipid peroxidation associated with lower mitochondrial membrane potential (Moore et al., 2023). Cyclic hypoxia in Mya arenaria increased proton leak and reduced OXPHOS coupling efficiency, consistent with oxidative membrane damage effects on mitochondrial coupling (Ouillon et al., 2021). In Lemna minor, lipid peroxidation preceded mitochondrial membrane potential reduction under gamma radiation and 3,5-dichlorophenol exposure, supporting this link in an aquatic primary producer (Xie et al., 2018; Xie et al., 2019). |
Direct studies measuring lipid peroxidation and OXPHOS coupling in the same exposure series are limited; mitochondrial dysfunction can also drive lipid peroxidation. |
|
Relationship 2203: decreased coupling of OXPHOS leads to decreased ATP pool |
High |
AOP 263 reports strong evidence for this KER (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). Cadmium exposure in oysters reduced state 3 respiration and affected mitochondrial bioenergetics (Sokolova et al., 2005). |
Compensatory glycolysis and altered metabolic demand can obscure total ATP changes. |
|
Relationship 2768: decreased ATP pool leads to increased cell injury/death |
Moderate to high |
ATP depletion and cell death are linked in multiple cell systems. Intracellular ATP concentration influences the decision between apoptosis and necrosis (Leist et al., 1997). Calcium electroporation caused dose-dependent ATP depletion and cancer cell death (Hansen et al., 2015). |
ATP assays may reflect both energy state and cell number; direct temporal separation of ATP depletion from cell death is needed. |
|
Relationship 2767: increased cell injury/death leads to decreased growth |
Moderate |
In Daphnia, toxicant-induced physiological energy disruption and cell/tissue injury are associated with growth reduction (Knops et al., 2001). In bivalves, cadmium and temperature interactions caused cellular energy disruption, mortality, and reduced condition/growth-related outcomes (Cherkasov et al., 2006). Methanol-exposed mouse embryos showed growth reduction and elevated cell death (Abbott et al., 1995). |
Growth can be reduced by mechanisms other than cell death; direct dose/time concordance between cell death and growth is not always measured. |
Inconsistencies and uncertainties
The main uncertainty for AOP 331 is the quantitative strength and directionality of the lipid peroxidation to OXPHOS coupling relationship. Lipid peroxidation can impair mitochondrial membranes, but mitochondrial dysfunction can also enhance ROS generation and thereby increase lipid peroxidation. AOP 331 represents one biologically plausible and empirically supported direction within a broader feedback-prone network. Another uncertainty is that ATP depletion can lead to different cellular outcomes depending on severity and duration; moderate depletion may reduce proliferation or activate adaptive stress responses, whereas severe depletion promotes cell injury/death. Finally, growth is a multifactorial endpoint. Increased cell injury/death is an important contributor to impaired growth, but decreased growth can also arise through reduced proliferation, altered cell size, altered energy allocation, endocrine signaling, or developmental delay without overt cell death.
Known Modulating Factors
|
Modulating factor |
Influence or outcome |
KER(s) involved |
|
Antioxidant capacity and redox buffering |
Higher antioxidant capacity reduces oxidative stress and lipid peroxidation; depletion or inhibition increases progression probability (Sies et al., 2017). |
2009, 3116 |
|
Membrane lipid composition and PUFA content |
High PUFA content increases susceptibility to lipid peroxidation and may increase effects on membrane potential and mitochondrial coupling (Ayala et al., 2014; Moore et al., 2023). |
3116, 1599 |
|
Mitochondrial density and metabolic demand |
Cells or taxa with high mitochondrial demand may be more susceptible to OXPHOS impairment, ATP depletion, and death (Murphy, 2009; Nicholls and Ferguson, 2013). |
1599, 2203, 2768 |
|
Oxygen availability and hypoxia-reoxygenation |
Fluctuating oxygen can increase ROS production, proton leak, mitochondrial impairment, and cell injury (Ouillon et al., 2021). |
2009, 3116, 1599 |
|
Temperature |
Temperature modifies membrane properties, oxygen demand, ROS production, mitochondrial performance, and growth. |
Multiple |
|
Compensatory energy metabolism |
Glycolysis and other ATP-generating pathways can buffer ATP depletion and delay injury/death. |
2203, 2768 |
|
Life stage and growth rate |
Rapidly growing or developing systems may be more sensitive to cell injury and loss of viable cell mass. |
2768, 2767 |
Quantitative Understanding
Quantitative understanding varies across the AOP. The relationship between OXPHOS coupling and ATP production has the strongest quantitative foundation, while the relationships linking oxidative stress to lipid peroxidation and cell injury/death to organismal growth are more often qualitative or semi-quantitative.
|
KER |
Quantitative understanding call |
Rationale |
|
2009: ROS increase to oxidative stress increase |
Low to moderate |
ROS measurements are reactive, transient, and assay-dependent. Quantitative relationships can be defined within a specific assay, but generalizable prediction across taxa and stressors remains limited (Sies et al., 2017). |
|
3116: oxidative stress increase to lipid peroxidation increase |
Low to moderate |
Dose-response relationships are reported for oxidative stress markers and lipid peroxidation, but lipid composition and assay differences strongly affect response magnitude (Ayala et al., 2014; Knauert and Knauer, 2008). |
|
1599: lipid peroxidation increase to decreased OXPHOS coupling |
Low to moderate |
Quantitative associations exist between lipid peroxidation and mitochondrial membrane potential or coupling efficiency, but broadly generalizable models are not established (Moore et al., 2023; Ouillon et al., 2021). |
|
2203: decreased OXPHOS coupling to decreased ATP pool |
High |
AOP 263 reports strong quantitative understanding, supported by bioenergetic theory and models linking mitochondrial coupling and ATP production (AOP-Wiki, 2026b; OECD, 2022; Song and Villeneuve, 2021). |
|
2768: decreased ATP pool to increased cell injury/death |
Moderate |
ATP thresholds influence the type and severity of cell death, and quantitative relationships are reported in defined systems, but thresholds vary by cell type and exposure condition (Leist et al., 1997; Hansen et al., 2015). |
|
2767: increased cell injury/death to decreased growth |
Low to moderate |
Quantitative linkage between cell loss and organismal growth is plausible and can be modeled in defined systems, but empirical cross-taxa response-response relationships remain limited (Conlon and Raff, 1999). |
BMD/POD-anchored concordance
The following benchmark-dose/point-of-departure (BMD/POD) concordance table anchors AOP 331 to quantitative cross-KE ordering, in line with Handbook section 4C. The multiomics point-of-departure (moPOD) dataset for gamma-irradiated Daphnia magna (Song et al., 2023) provides POD magnitudes for increased ROS, decreased ATP, decreased OXPHOS coupling, and cell death, demonstrating the expected upstream-to-downstream POD ordering (more sensitive PODs upstream). The moPOD is presented as POD magnitude evidence, not as a causal re-ordering of KEs. The Lemna minor EDR50 range provides a whole-pathway apical anchor in an aquatic primary producer.
|
Key event (functional category) |
POD metric |
POD value (mGy/h) |
POD ordering |
Source |
|
KE 1115: ROS, increased (mROS) |
moPOD (multiomics POD) |
0.4 |
1 (most sensitive) |
Song et al., 2023 |
|
KE 1771: ATP pool, decreased |
moPOD |
2.5 |
2 |
Song et al., 2023 |
|
KE 1446: OXPHOS coupling, decreased (UPS/OXPHOS module) |
moPOD |
42.3 |
3 |
Song et al., 2023 |
|
KE 55: Cell injury/death (apoptosis) |
moPOD |
42.3 |
3 (least sensitive) |
Song et al., 2023 |
|
Upstream KE chain → growth (Lemna minor, gamma) |
EDR50 (growth) |
31.5–54.8 (mGy/h) |
whole-pathway apical |
Xie et al., 2018, 2019, 2022 |
Considerations for Potential Applications of the AOP (optional)
AOP 331 can support mechanistic interpretation of growth impairment caused by oxidative stressors that induce lipid peroxidation, mitochondrial bioenergetic dysfunction, ATP depletion, and cell injury/death. The AOP is particularly relevant for hazard identification and chemical prioritization when evidence indicates increased ROS or oxidative stress together with lipid peroxidation, mitochondrial membrane potential changes, reduced respiratory control, ATP depletion, cytotoxicity, or growth inhibition. The AOP may also support IATA development by linking upstream NAM endpoints, such as ROS assays, lipid peroxidation markers, mitochondrial membrane potential, oxygen consumption rate, ATP content, cytotoxicity assays, and organismal growth measurements.
AOP 331 can support chemical grouping and read-across for stressors that share evidence of oxidative lipid damage, mitochondrial bioenergetic impairment, and ATP-associated cell injury. Because oxidative stress and lipid peroxidation are not chemical-specific, this AOP should not be used as a stand-alone basis for regulatory decisions. Instead, it should be applied as part of a weight-of-evidence framework that considers stressor mode of action, exposure context, assay specificity, taxonomic relevance, and concordance across multiple KEs. The AOP also highlights method-development needs, particularly standardized assays for lipid peroxidation, OXPHOS coupling, ATP depletion, and cell injury/death endpoints that can be connected quantitatively to apical growth outcomes.
References
Abbott BD, Harris MW, Birnbaum LS. 1995. Cell death in rat and mouse embryos exposed to methanol in whole embryo culture: evaluation of the role of the p53 tumor suppressor gene. Teratogenesis, Carcinogenesis, and Mutagenesis 15:147-169.
AOP-Wiki. 2026a. AOP 478: Deposition of energy leading to occurrence of cataracts. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/478.
AOP-Wiki. 2026b. AOP 263: Uncoupling of oxidative phosphorylation leading to growth inhibition via decreased cell proliferation. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/263.
AOP-Wiki. 2026c. AOP 12: Chronic binding of antagonist to N-methyl-D-aspartate receptors during brain development induces impairment of learning and memory abilities. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/12.
AOP-Wiki. 2026d. AOP 13: Chronic binding of antagonist to N-methyl-D-aspartate receptors during brain development induces impairment of learning and memory abilities. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/13.
AOP-Wiki. 2026e. AOP 17: Binding of electrophilic chemicals to SH/seleno-proteins involved in protection against oxidative stress leading to impairment of learning and memory. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/17.
AOP-Wiki. 2026f. AOP 38: Protein alkylation leading to liver fibrosis. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/38.
AOP-Wiki. 2026g. AOP 48: Binding of agonists to ionotropic glutamate receptors in adult brain causes excitotoxicity that mediates neuronal cell death, contributing to learning and memory impairment. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/48.
AOP-Wiki. 2026h. AOP 296: Oxidative DNA damage leading to chromosomal aberrations and mutations. Collaborative Adverse Outcome Pathway Wiki. Available from: https://aopwiki.org/aops/296.
Ayala A, Munoz MF, Arguelles S. 2014. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Medicine and Cellular Longevity 2014:360438.
Barata C, Varo I, Navarro JC, Arun S, Porte C. 2005. Antioxidant enzyme activities and lipid peroxidation in the freshwater cladoceran Daphnia magna exposed to redox cycling compounds. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 140:175-186.
Bonora M, Bononi A, De Marchi E, Giorgi C, Lebiedzinska M, Marchi S, Patergnani S, Rimessi A, Suski JM, Wojtala A, Wieckowski MR, Kroemer G, Galluzzi L, Pinton P. 2012. Role of the c subunit of the FO ATP synthase in mitochondrial permeability transition. Cell Cycle 12:674-683.
Carvaillo JC, Barouki R, Coumoul X, Audouze K. 2019. Linking bisphenol S to adverse outcome pathways using a combined text mining and systems biology approach. Environmental Health Perspectives 127:047005.
Cherkasov AS, Biswas PK, Ridings DM, Ringwood AH, Sokolova IM. 2006. Effects of acclimation temperature and cadmium exposure on cellular energy budgets in the marine mollusk Crassostrea virginica: linking cellular and mitochondrial responses. Journal of Experimental Biology 209:1274-1284.
Conlon I, Raff M. 1999. Size control in animal development. Cell 96:235-244.
Esperanza M, Cid A, Herrero C, Rioboo C. 2015. Acute effects of a prooxidant herbicide on the microalga Chlamydomonas reinhardtii: screening cytotoxicity and genotoxicity endpoints. Aquatic Toxicology 165:210-221.
Hansen EL, Sozer EB, Romeo S, Frandsen SK, Vernier PT, Gehl J. 2015. Dose-dependent ATP depletion and cancer cell death following calcium electroporation, relative effect of calcium concentration and electric field strength. PLoS ONE 10:e0122973.
Jornod F, Jaylet T, Blaha L, Sarigiannis D, Tamisier L, Audouze K. 2022. AOP-helpFinder webserver: a tool for comprehensive analysis of the literature to support adverse outcome pathways development. Bioinformatics 38:1173-1175.
Knauert S, Knauer K. 2008. The role of reactive oxygen species in copper toxicity to two freshwater green microalgae. Journal of Phycology 44:311-321.
Knops M, Altenburger R, Segner H. 2001. Alterations of physiological energetics, growth and reproduction of Daphnia magna under toxicant stress. Aquatic Toxicology 53:79-90.
Leist M, Single B, Castoldi AF, Kuhnle S, Nicotera P. 1997. Intracellular adenosine triphosphate concentration: a switch in the decision between apoptosis and necrosis. Journal of Experimental Medicine 185:1481-1486.
Moore TD, Martin-Creuzburg D, Yampolsky LY. 2023. Diet effects on longevity, heat tolerance, lipid peroxidation and mitochondrial membrane potential in Daphnia. Oecologia 202:151-163.
Murphy MP. 2009. How mitochondria produce reactive oxygen species. Biochemical Journal 417:1-13.
Nicholls DG, Ferguson SJ. 2013. Bioenergetics 4. London: Academic Press.
Nicotera P, Leist M, Ferrando-May E. 1998. Intracellular ATP, a switch in the decision between apoptosis and necrosis. Toxicology Letters 102-103:139-142. https://doi.org/10.1016/S0378-4274(98)00298-7
OECD. 2018. Users' handbook supplement to the guidance document for developing and assessing adverse outcome pathways. OECD Series on Adverse Outcome Pathways No. 1. Paris: OECD Publishing.
OECD. 2021. Guidance document for the scientific review of adverse outcome pathways. OECD Series on Testing and Assessment No. 344. Paris: OECD Publishing.
OECD. 2022. Uncoupling of oxidative phosphorylation leading to growth inhibition via decreased cell proliferation. OECD Series on Adverse Outcome Pathways No. 28. Paris: OECD Publishing.
Ouillon N, Sokolov EP, Otto S, Rehder G, Sokolova IM. 2021. Effects of variable oxygen regimes on mitochondrial bioenergetics and reactive oxygen species production in a marine bivalve, Mya arenaria. Journal of Experimental Biology 224:jeb237156.
Qian H, Chen W, Sun L, Jin Y, Liu W, Fu Z. 2009. Inhibitory effects of paraquat on photosynthesis and the response to oxidative stress in Chlorella vulgaris. Ecotoxicology 18:537-543.
Schieber M, Chandel NS. 2014. ROS function in redox signaling and oxidative stress. Current Biology 24:R453-R462.
Sies H, Berndt C, Jones DP. 2017. Oxidative stress. Annual Review of Biochemistry 86:715-748.
Sokolova IM, Sokolov EP, Ponnappa KM. 2005. Cadmium exposure affects mitochondrial bioenergetics and gene expression of key mitochondrial proteins in the eastern oyster Crassostrea virginica Gmelin (Bivalvia: Ostreidae). Aquatic Toxicology 73:242-255.
Song Y, Villeneuve DL. 2021. Uncoupling of oxidative phosphorylation leading to growth inhibition via decreased cell proliferation. Environmental Toxicology and Chemistry 40:2951-2963.
Xie L, Gomes T, Solhaug KA, Song Y, Tollefsen KE. 2018. Linking mode of action of the model respiratory and photosynthesis uncoupler 3,5-dichlorophenol to adverse outcomes in Lemna minor. Aquatic Toxicology 197:98-108. https://doi.org/10.1016/j.aquatox.2018.02.005
Xie L, Solhaug KA, Song Y, Brede DA, Lind OC, Salbu B, Tollefsen KE. 2019. Modes of action and adverse effects of gamma radiation in an aquatic macrophyte Lemna minor. Science of the Total Environment 680:23-34. https://doi.org/10.1016/j.scitotenv.2019.05.016
Xie L, Song Y, Petersen K, Solhaug KA, Lind OC, Brede DA, Salbu B, Tollefsen KE. 2022. Ultraviolet B modulates gamma radiation-induced stress responses in Lemna minor at multiple levels of biological organisation. Science of the Total Environment 846:157457. https://doi.org/10.1016/j.scitotenv.2022.157457