
This AOP is licensed under the BY-SA license. This license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 323
Title
PPARalpha Agonism Leading to Decreased Viable Offspring via Decreased 11-Ketotestosterone
Short name
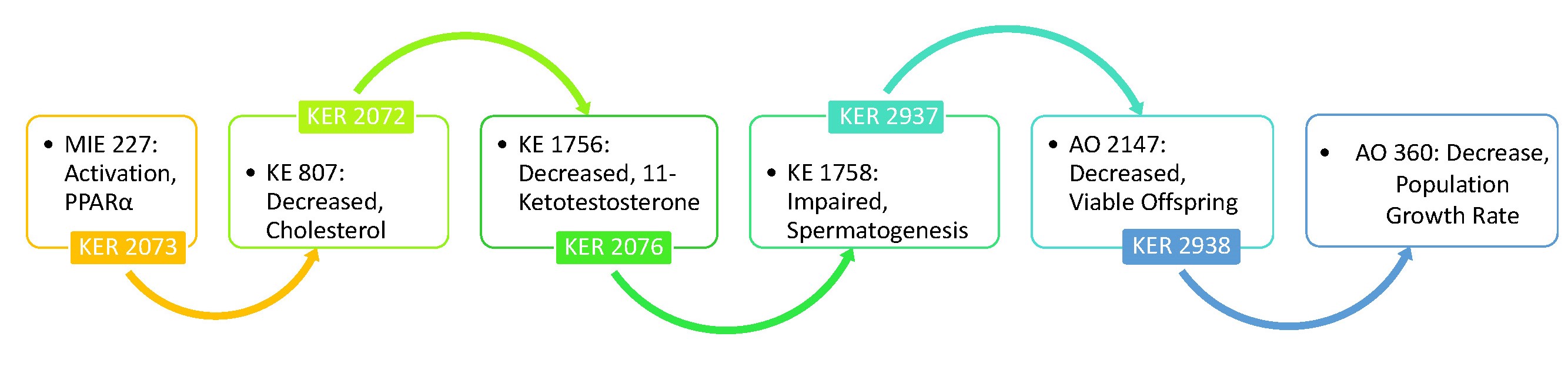
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 323, PPARalpha Agonism Leading to Decreased Viable Offspring via Decreased 11-Ketotestosterone in tools offered by third parties.
Point of Contact
Contributors
- Ashley Kittelson
- John Hoang
- Robin Kutsi
- Jennifer Olker
Coaches
- Dan Villeneuve
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on September 28, 2023 16:35
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Activation, PPARα | December 28, 2020 12:48 |
| Decreased, cholesterol | May 24, 2022 11:10 |
| Decreased, plasma 11-ketotestosterone level | May 24, 2022 13:51 |
| Impaired, Spermatogenesis | July 28, 2026 10:54 |
| Decrease, Population growth rate | January 03, 2023 09:09 |
| Decreased, Viable Offspring | April 10, 2024 17:43 |
| Activation, PPARα leads to Decreased, cholesterol | May 01, 2020 11:10 |
| Decreased, cholesterol leads to Decreased, 11KT | May 06, 2020 09:31 |
| Decreased, 11KT leads to Impaired, Spermatogenesis | April 19, 2021 13:32 |
| Impaired, Spermatogenesis leads to Decreased, Viable Offspring | September 29, 2023 12:58 |
| Decreased, Viable Offspring leads to Decrease, Population growth rate | September 29, 2023 10:57 |
| Clofibrate | November 29, 2016 18:42 |
| Gemfibrozil | March 31, 2020 10:24 |
| Fenofibrate | November 29, 2016 18:42 |
Abstract
This adverse outcome pathway details the linkage from peroxisome proliferator-activated receptor alpha (PPARα) activation to the adverse effects of decreased viable offspring and decrease in population growth rate in fish. PPARα is a ligand-activated nuclear receptor that, after forming a heterodimer with retinoid X receptor (RXR), promotes transcription of many genes including those involved in fatty acid β-oxidation and cholesterol metabolism. Synthetic ligands have been designed as pharmaceuticals to target PPARα for treatment of human metabolic diseases. Exposure to these pharmaceuticals or other contaminants in environment can disrupt metabolic processes in fish, including the activation of PPARα. In fish, this can lead to decreased cholesterol which in turn causes a decrease in reproductive hormones, notably 11-ketotestosterone (11-KT). A decrease in reproductive hormones impairs the fish’s ability to reproduce. Described here is the pathway in which decreased 11-KT impairs inducement of spermatogenesis and sperm production which results in a reduced number of viable offspring. This can lead to impacts on population growth rate due to the decreased number of viable offspring resulting in a decline in recruitment and contribution of offspring to the next generation.
AOP Development Strategy
Context
This AOP was developed to address one potential effect of per- and polyfluoroalkyl substances (PFAS) on fish. Through review of the human health and in vitro toxicity data on conserved pathways and molecular targets for PFAS disruption, activation of PPARα was identified as a potential target of several PFAS which could result in altered lipid metabolism. This AOP focused primarily on teleost fish using experimental data from prototypical stressors, along with knock-out and genetic mutation experiments, for evidence of causality and essentiality for existing and newly developed KEs and KERs.
Strategy
Biologists under the guidance of experts in the field developed this AOP with a KER by KER approach. This collaborative effort included dividing the KERs for investigation and evaluation, with regular team meetings to review findings with leaders in AOP development and fish toxicity testing (Dr. Daniel Villeneuve, Dr. Gerald Ankley, Kathleen Jensen). Systematic approaches were incorporated into the methods for identification and review of relevant studies for increased transparency and reproducibility. Literature reviews for fibrates (synthetic ligands for PPARα) were conducted to develop and support the early KERs (PPARα activation through impaired spermatogenesis) whereas the intermediate KERs were supported with empirical evidence from published fish literature on prototypical stressors and hormones that disrupt the endocrine system. Additional sources were reviewed and incorporated based on citation tracing and expert knowledge. The resulting concordance tables and key supporting papers underwent a secondary quality assurance review by a team member (Kathleen Jensen) with extensive experience in fish toxicity testing and endocrinology.
All of the KERs were newly developed for this AOP, using an existing MIE (Activation, PPARα), one existing KE (Decreased, cholesterol), and novel KEs for the remaining steps through the organism level adverse outcome (Decreased, viable offspring). The population level adverse outcome (Decrease, population growth rate) was already relevant for this AOP and did not need to be updated.
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 227 | Activation, PPARα | Activation, PPARα |
| KE | 807 | Decreased, cholesterol | Decreased, cholesterol |
| KE | 1756 | Decreased, plasma 11-ketotestosterone level | Decreased, 11KT |
| KE | 1758 | Impaired, Spermatogenesis | Impaired, Spermatogenesis |
| AO | 2147 | Decreased, Viable Offspring | Decreased, Viable Offspring |
| AO | 360 | Decrease, Population growth rate | Decrease, Population growth rate |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Activation, PPARα leads to Decreased, cholesterol | adjacent | High | Low |
| Decreased, cholesterol leads to Decreased, 11KT | adjacent | High | Low |
| Decreased, 11KT leads to Impaired, Spermatogenesis | adjacent | High | Low |
| Impaired, Spermatogenesis leads to Decreased, Viable Offspring | adjacent | Moderate | Low |
| Decreased, Viable Offspring leads to Decrease, Population growth rate | adjacent | Moderate | Low |
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Adult | High |
Taxonomic Applicability
| Term | Scientific Term | Evidence | Link |
|---|---|---|---|
| teleost fish | teleost fish | High | NCBI |
Sex Applicability
| Sex | Evidence |
|---|---|
| Male | High |
Overall Assessment of the AOP

Domain of Applicability
The empirical evidence suggests that this AOP is applicable to adult, reproductively mature, male teleost fish.
Life Stage
The life stage applicable to this AOP is adult, reproductively mature organisms.
Sex
The process of spermatogenesis occurs in reproductively mature males. Therefore, this AOP is only applicable to males.
Taxonomic
This AOP is considered most relevant for teleost fish. Most of the experimental evidence compiled for this AOP is from teleost fish, for which 11-KT is the dominant androgen. However, PPARs including PPARα are highly conserved across humans, rodents, and fish. An evaluation of protein sequence conservation via SeqAPASS (https://seqapass.epa.gov/seqapass/) predicted similarity in cross-species susceptibility to PPARα agonists among humans, zebrafish, medaka, and other fish species. Thus, PPARα agonism and downstream effects on cholesterol, hormone production (not limited to 11-KT), spermatogenesis (a highly conserved biological process), and production of offspring could have more broad taxonomic relevance.
Essentiality of the Key Events
Essentiality of most of key events in this AOP is supported with experimental exposures with prototypical stressors that demonstrate modification of a more upstream KE associated with a corresponding change in downstream KE(s). Several of the key events have further support for essentiality with knock-out and genetic mutations experiments as well as rescue studies. Key studies are listed below.
Although it is challenging to directly measure PPARα activation in fish in vivo studies, there are multiple studies that have shown that fish exposed to fibrates (and thus assumed activation of PPARα) have decreased cholesterol. This relationship has been demonstrated in a variety of fish species [fathead minnow (Runnalls et al., 2007), grass carp (Du et al., 2008; Guo et al., 2015), Nile tilapia (Ning et al., 2017), rainbow trout (Prindiville et al., 2011), medaka (Lee et al., 2019), zebrafish (AL-Habsi et al., 2016; Velasco-Santamaria et al., 2011; Fraz et al., 2018), turbot (Urbatzka et al., 2015)], with temporal and dose concordance in one study (Velasco-Santamaria et al., 2011).
The process of steroid hormone biosynthesis is well understood, and cholesterol is the precursor for all steroid hormones, including 11-KT. The relationship between decreased cholesterol and decreased 11-KT is well-established. There are several experimental exposure studies that showed decreased 11-KT associated with decreased cholesterol with dose and temporal concordance (Lee et al., 2019; Velasco-Santamaria et al., 2011). The essentiality of cholesterol for production of 11-KT is further supported by an ex vivo study which showed that exposure to gemfibrozil (a known PPARα agonist) resulted in decreased 11-KT production unless supplemented with 25OH-cholesterol (Fraz et al., 2018), demonstrating that decreased cholesterol availability was the cause of the decreased steroid synthesis.
11-KT is well documented as a critical androgen for proper male reproduction in teleost fish and has well-documented involvement in spermatogenesis and spermiation. The essentiality of 11-KT for spermatogenesis has been documented in zebrafish knock-out studies with rescue (Zhang et al., 2020) which showed that zebrafish with cyp11c1 knockout have reduced 11-KT levels, smaller genitalia, inability naturally mate, defective Leydig and Sertoli cells, and insufficient spermatogenesis. The treatment of100 nM 11-KA (which is converted to 11-KT in vivo) for 4 hours per day for 10 days corrected these effects, demonstrating that insufficient 11-KT levels was the cause of arrested spermatogenesis.
Successful oocyte fertilization and production of viable offspring is dependent on spermatogenesis and the production of sufficient quality and quantity of sperm. Essentiality is strongly supported by gene modification studies, such knock-out studies targeting genes associated with spermatogenesis and meiotic division as well as exposure studies with known endocrine disruptors (e.g., DEHP, EE2). Multiple studies with zebrafish have shown that knockouts targeting genes associated with spermatogenesis (e.g., Tdrd12, AR) and meiotic division (e.g., E2f5, Mettl3, mlh1) resulted in interference with spermatogenesis (i.e., delayed or arrested progression, apoptosis, and decrease in sperm density, quality and/or motility) and male zebrafish that were either infertile or exhibited decreased fertilization rates when mated with WT females (Dai et al., 2017; Leal et al, 2008; Tang et al., 2018; Xia et al., 2018; Xie et al., 2020).
By definition, there must be viable offspring to maintain a population. However, there are other vital rates that are essential here as well, such as survival to reproductive age.
Evidence Assessment
The weight of evidence for each of the KERs within this AOP are ranked moderate to high. Each of the KERs is biologically plausible, with the highest ratings for the intermediate KERs (Decreased, cholesterol leading to Decreased, 11-KT and Decreased, 11-KT leading to Impaired, Spermatogenesis). The relationship between the MIE and the first key event is considered moderate for biological plausibility due to challenges in directly measuring the PPARα activation in in vivo studies. Whereas the links to the individual adverse outcome (Decreased, viable offspring) and the population level adverse outcome are considered moderate for biological plausibility due to the other factors that can influence each of these outcomes. There is substantial experimental evidence in fish to support this AOP, however, few studies measured multiple sequential key events and the final link to decreased population growth rate is based on biological plausibility and population modeling. Overall weight of evidence is moderate.
Biological Plausibility
This AOP is considered highly plausible, based on the evaluation of available evidence for the mechanistic (structural or functional) relationships between upstream and downstream KEs that are consistent with established biological knowledge. There is a broad understanding of lipid metabolism pathways and supporting in vivo and in vitro experimental data on the role of PPARα in lipid metabolism. PPARα is conserved across vertebrates and has been documented in multiple fish species, therefore biological plausibility is considered moderate for activation of PPARα leading to decreased cholesterol. The next two KERs are considered highly plausible. The process of steroid hormone biosynthesis is well understood, and cholesterol is the precursor for all steroid hormones including testosterone and 11-ketotestosterone (Norris and Carr, 2020). Similarly, the 11-ketotestosterone is well documented as necessary for spermatogenesis and sperm production (Amer et al., 2001; Borg, 1994; Geraudie et al., 2010). Because there are multiple factors required to produce viable offspring, biological plausibility is considered moderate for the process of impaired spermatogenesis leading to decreased viable offspring. The link from the individual level adverse outcome (decreased viable offspring) to the population level adverse outcome (decrease in population growth rate) is also influenced by multiple factors.
Empirical Support
There is substantial experimental evidence to support this AOP. Experimental results from a variety of fish studies with prototypical stressors demonstrate concordance and consistency throughout the AOP. However, there were few studies that measured multiple sequential KEs and limited concentration or dose-response data and temporal measurements across diversity of taxa. Due to these limitations, response-response relationships for a quantitative understanding of this AOP could not be evaluated. Concordance of empirical support across the AOP is summarized in Attachment A.
There are multiple studies in fish that demonstrate exposure to known PPARα agonists (considered prototypical stressors or model chemicals) resulted in decreased total cholesterol. These studies include experimental exposure of seven different fish species [fathead minnow (Runnalls et al., 2007), grass carp (Du et al., 2008; Guo et al., 2015), Nile tilapia (Ning et al., 2017), rainbow trout (Prindiville et al., 2011), medaka (Lee et al., 2019), zebrafish (AL-Habsi et al., 2016; Velasco-Santamaria et al., 2011; Fraz et al., 2018), turbot (Urbatzka et al., 2015)] to several different fibrates (clofibrate, clofibric acid, gemfibrozil, fenofibrate, WY-14643). Temporal and dose concordance was demonstrated in one study ( Velasco-Santamaría et al., 2011); however, there is insufficient empirical evidence for development of a quantitative relationship between the KEs.
While the following KER (decreased cholesterol leading to decreased 11-KT) has strong biological plausibility, there are relatively few fish experimental exposure studies that measured both cholesterol and 11-KT. Two exposure studies that measured both KEs showed dose and temporal concordance (Lee et al., 2019; Velasco-Santamaria et al., 2011), and the third study provided strong evidence essentiality of cholesterol for the production of 11-KT (Fraz et al., 2018).
There is substantial empirical evidence showing spermatogenesis in numerous fish species is dependent on 11-KT, with several studies demonstrating temporal and dose concordance for this relationship. These studies include testing of both higher 11-KT (treatments with 11-KT or increased production) and decreased 11-KT. For example, increased 11-KT has been related to measures of successful spermatogenesis such as greater number of spermatids (Agulleiro et al., 2007; Selvaraj et al., 2013), more advanced testicular stages (Cavaco et al., 1998, 2001), and more differentiated and later type spermatogonia (Melo et al., 2015; Miura et al., 1991). Whereas, decreased 11-KT in fish has been associated with negative impacts or delays in spermatogenesis including decreased number of spermatocytes, spermatids, and/or spermatozoa (Agbohessi et al., 2015; Chen et al., 2017; de Waal et al., 2009; Liu et al., 2018; Pereira et al., 2015; Sales et al., 2020; Xia et al,. 2018). Melo et al. (2015) is one example of studies that demonstrated temporal concordance; in this study exposure to adrenosterone (ketoandrostenedione; which is converted to 11-KT in vivo) caused an increase in 11-KT levels at 7 and 14 d, with Type A differentiated spermatogonial numbers also increased 14 d after treatment.
There is substantial empirical evidence demonstrating that impaired spermatogenesis results in decreased oocyte fertilization and a reduction in viable offspring. Much of the cited literature is from fish exposed to prototypical stressors (endocrine disruptors), with several studies demonstrating dose and temporal concordance. In addition to the gene modification studies previously described for essentiality, exposure studies with endocrine disruptors [e.g., di(2-ethylhexyl) phthalate (DEHP), 17α-ethinylestradiol (EE2), nonylphenol] provide evidence of concordance and consistency of this KER. These include studies with zebrafish, Nile tilapia, Japanese medaka, and marine medaka (Corradetti et al., 2013; Hill & Janz, 2003; Kang et al., 2002, Nash et al., 2004; Seki et al., 2002). Several studies provide evidence of dose-response concordance such as a concentration dependent effect on both spermatogenesis and fertilization rate of when male fish exposed to DEHP are mated with wild-type females (Ma et al., 2018; Uren-Webster et al., 2010; Ye et al., 2014).
Direct empirical evidence on population size decreases associated with decreased viable offspring is very limited. There are no empirical data suitable for evaluating the dose-response, temporal, or incidence concordance between these two adverse outcomes. This relationship is based on biological plausibility and population modeling (e.g., Miller & Ankley, 2004; Miller et al., 2020).
Uncertainties, inconsistencies, and data gaps
- There were no notable inconsistencies in the literature that was reviewed for development of this AOP. However, there are several areas of uncertainty. These include:
- It is challenging to directly measure PPARα agonism in fish in vivo studies. Therefore, we relied on fish exposure studies with pharmaceuticals designed to activate PPARα in humans. However, there is uncertainty of whether all fibrates shown effective in humans are PPARα agonists in fish. This AOP was developed on the assumption that these pharmaceutical also activate PPARα in fish, which is supported by a cross-species comparison in vitro and susceptibility evaluation based on gene sequences support similarity in responses across vertebrates.
- 11-KT levels can be highly variable between fish species and have seasonal fluctuations within a species (with highest levels at spawning).
- For the relationship between 11-KT and spermatogenesis, a few studies documented a significant change in one without a significant change in the other, highlighting the complexity of this relationship.
- Both of the adverse outcomes (Decreased, viable offspring and Decrease, population growth rate) are influenced by multiple factors. The key events in this AOP are just one potential path to these outcomes. In addition, PPARα agonism could result in other toxicity pathways, such as decreased juvenile growth, which were not included in the development of this AOP.
- Finally, few studies measured multiple sequential key events; thus evidence had to be compiled KER by KER to support this AOP.
Known Modulating Factors
Quantitative Understanding
At this time available data are insufficient to develop a quantitative AOP linking PPARα agonism with decreased viable offspring or decreased population growth rate.
Considerations for Potential Applications of the AOP (optional)
- The present AOP can inform a tiered testing approach for PPARα agonists (including some PFAS) based on in vitro screening results (e.g., Houck et al., 2021) and targeted in vivo testing (illustrated by Villeneuve et al., 2023).
- The present AOP can inform the development of microphysiological or computational systems models to evaluated probable effects on reproduction.
- The present AOP can aid in prediction of potential effects when PPAR agonists are measured in environmental samples and interpretation (along with selection of additional endpoints to measure) when PPAR activity is detected with effects-based environmental monitoring (e.g., Blackwell et al., 2019).
References
Agbohessi, P.T., Imorou Toko I., Ouédraogo, A., Jauniaux, T., Mandiki, S.N., & Kestemont, P. (2015). Assessment of the health status of wild fish inhabiting a cotton basin heavily impacted by pesticides in Benin (West Africa). Science of the Total Environment, 506-507, 567-584. https://doi.org/10.1016/j.scitotenv.2014.11.047
Agulleiro, M.J., Scott, A.P., Duncan, N., Mylonas, C.C., & Cerdà, J. (2007). Treatment of GnRHa-implanted Senegalese sole (Solea senegalensis) with 11-ketoandrostenedione stimulates spermatogenesis and increases sperm motility. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 147(4), 885-92. https://doi.org/10.1016/j.cbpa.2007.02.008
Al-Habsi, A.A., A. Massarsky, T.W. Moon (2016) “Exposure to gemfibrozil and atorvastatin affects cholesterol metabolism and steroid production in zebrafish (Danio rerio)”, Comparative Biochemistry and Physiology, Part B, Vol. 199, Elsevier, pp. 87-96. http://dx.doi.org/10.1016/j.cbpb.2015.11.009
Amer, M.A., Miura, T., Miura, C., & Yamauchi, K. (2001). Involvement of Sex Steroid Hormones in the Early Stages of Spermatogenesis in Japanese Huchen (Hucho perryi ). Biology of Reproduction, 65(4), 1057–1066. https://doi.org/10.1095/biolreprod65.4.1057
Blackwell, B. R., Ankley, G. T., Bradley, P. M., Houck, K. A., Makarov, S. S., Medvedev, A. V., Swintek, J., & Villeneuve, D. L. (2019). Potential Toxicity of Complex Mixtures in Surface Waters from a Nationwide Survey of United States Streams: Identifying in Vitro Bioactivities and Causative Chemicals. Environmental science & technology, 53(2), 973–983. https://doi.org/10.1021/acs.est.8b05304
Borg, B. (1994). Androgens in teleost fishes. Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology, and Endocrinology, 109(3), 219-245. https://doi.org/10.1016/0742-8413(94)00063-G
Cavaco, J.E.B., Bogerd, J., Goos, H., & Schulz, R.W. (2001). Testosterone inhibits 11-ketotestosterone-induced spermatogenesis in African catfish (Clarias gariepinus). Biology of Reproduction, 65(6), 1807-1812. https://doi.org/10.1095/biolreprod65.6.1807
Cavaco, J.E.B., Vilrokx, C., Trudeau, V.L., Schulz, R.W., & Goos, H.J.T. (1998). Sex steroids and the initiation of puberty in male African catfish, Clarias gariepinus. American Journal of Physiology, 275(6), 1793-1802. https://doi.org/10.1152/ajpregu.1998.275.6.R1793
Chen, J., Jiang, D., Tan, D., Fan, Z., Wei, Y., Li, M., & Wang, D. (2017). Heterozygous mutation of eEF1A1b resulted in spermatogenesis arrest and infertility in male tilapia, Oreochromis niloticus. Scientific Reports, 7, 43733. https://doi.org/10.1038/srep43733
Corradetti, B., Stronati, A., Tosti, L., Manicardi, G., Carnevali, O., & Bizzaro, D. (2013). Bis-(2-ethylexhyl) phthalate impairs spermatogenesis in zebrafish (Danio rerio). Reproductive biology, 13(3), 195–202. https://doi.org/10.1016/j.repbio.2013.07.003
Dai, X., Shu, Y., Lou, Q., Tian, Q., Zhai, G., Song, J., Lu, S., Yu, H., He, J., & Yin, Z. (2017). Tdrd12 Is Essential for Germ Cell Development and Maintenance in Zebrafish. International journal of molecular sciences, 18(6), 1127. https://doi.org/10.3390/ijms18061127 de Waal, P.P., Leal, M.C., García-López, A., Liarte, S., de Jonge, H., Hinfray, N., Brion, F., Schulz, R.W., & Bogerd, J. (2009). Oestrogen-induced androgen insufficiency results in a reduction of proliferation and differentiation of spermatogonia in the zebrafish testis. Journal of Endocrinology, 202(2), 287-97. https://doi.org/10.1677/JOE-09-0050
Du, Z. et al. (2008) “Hypolipidaemic effects of fenofibrate and fasting in the herbivorous grass carp (Ctenopharyngodon Idella) fed a high-fat diet”, British Journal of Nutrition, Vol. 100, Cambridge University Press, pp. 1200-1212. doi:10.1017/S0007114508986840
Fraz, S., A.H. Lee, J.Y. Wilson (2018) “Gemfibrozil and carbamazepine decrease steroid production in zebrafish testes (Danio rerio)”, Aquatic Toxicology, Vol. 198, Elsevier, pp. 1-9. https://doi.org/10.1016/j.aquatox.2018.02.006
Geraudie, P., Gerbron, M., & Minier, C. (2010). Seasonal variations and alterations of sex steroid levels during the reproductive cycle of male roach (Rutilus rutilus). Marine Environmental Research, 69(S1), S53-S55. https://doi.org/10.1016/j.marenvres.2009.11.008
Guo, X. et al. (2015) “Effects of lipid-lowering pharmaceutical clofibrate on lipid and lipoprotein metabolism of grass carp (Ctenopharyngodon idella Val.) fed with the high non-protein energy diets”, Fish Physiology and Biochemistry, Vol. 41, Springer, pp. 331-343. doi: 10.1007/s10695-014-9986-8
Hill, R. L., Jr, & Janz, D. M. (2003). Developmental estrogenic exposure in zebrafish (Danio rerio): I. Effects on sex ratio and breeding success. Aquatic toxicology (Amsterdam, Netherlands), 63(4), 417–429. https://doi.org/10.1016/s0166-445x(02)00207-2
Houck, K. A., Patlewicz, G., Richard, A. M., Williams, A. J., Shobair, M. A., Smeltz, M., Clifton, M. S., Wetmore, B., Medvedev, A., & Makarov, S. (2021). Bioactivity profiling of per- and polyfluoroalkyl substances (PFAS) identifies potential toxicity pathways related to molecular structure. Toxicology, 457, 152789. https://doi.org/10.1016/j.tox.2021.152789
Kang, I. J., Yokota, H., Oshima, Y., Tsuruda, Y., Yamaguchi, T., Maeda, M., Imada, N., Tadokoro, H., & Honjo, T. (2002). Effect of 17beta-estradiol on the reproduction of Japanese medaka (Oryzias latipes). Chemosphere, 47(1), 71–80. https://doi.org/10.1016/s0045-6535(01)00205-3
Leal, M. C., Feitsma, H., Cuppen, E., França, L. R., & Schulz, R. W. (2008). Completion of meiosis in male zebrafish (Danio rerio) despite lack of DNA mismatch repair gene mlh1. Cell and tissue research, 332(1), 133–139. https://doi.org/10.1007/s00441-007-0550-z
Lee, G. et al. (2019) Effects of gemfibrozil on sex hormones and reproduction related performances of Oryzias latipes following long-term (155 d) and short-term (21 d) exposure, Ecotoxicology and Environmental Safety, Vol. 173, Elsevier, pp. 174-181. https://doi.org/10.1016/j.ecoenv.2019.02.015
Liu, Z.H., Chen, Q.L., Chen, Q., Li, F., & Li, Y.W. (2018). Diethylstilbestrol arrested spermatogenesis and somatic growth in the juveniles of yellow catfish (Pelteobagrus fulvidraco), a fish with sexual dimorphic growth. Fish Physiology and Biochemistry, 44(3), 789-803. https://doi.org/10.1007/s10695-018-0469-1
Ma, Yan-Bo, Jia, Pan-Pan, Junaid, Muhammad, Yang, Li, Lu, Chun-Jiao, & Pei, De-Sheng. (2018). Reproductive effects linked to DNA methylation in male zebrafish chronically exposed to environmentally relevant concentrations of di-(2-ethylhexyl) phthalate. Environmental Pollution (1987), 237, 1050-1061.
Melo, M.C., van Dijk, P., Andersson, E., Nilsen, T.O., Fjelldal, P.G., Male, R., Nijenhuis, W., Bogerd, J., de França, L.R., Taranger, G.L., & Schulz R.W. (2015). Androgens directly stimulate spermatogonial differentiation in juvenile Atlantic salmon (Salmo sala). General and Comparative Endocrinology, 211, 52-61. https://doi.org/10.1016/j.ygcen.2014.11.015.
Miller, D. H., & Ankley, G. T. (2004). Modeling impacts on populations: fathead minnow (Pimephales promelas) exposure to the endocrine disruptor 17beta-trenbolone as a case study. Ecotoxicology and environmental safety, 59(1), 1–9. https://doi.org/10.1016/j.ecoenv.2004.05.005
Miller, D. H., Clark, B. W., & Nacci, D. E. (2020). A multidimensional density dependent matrix population model for assessing risk of stressors to fish populations. Ecotoxicology and environmental safety, 201, 110786. https://doi.org/10.1016/j.ecoenv.2020.110786
Miura, T., Yamauchi, K., Takahashi, H., & Nagahama, Y. (1991). Hormonal induction of all stages of spermatogenesis in vitro in the male Japanese eel (Anguilla japonica). Proceedings of the National Academy of Sciences of the United States of America, 88(13), 5774–5778. https://doi.org/10.1073/pnas.88.13.5774
Ning, L. et al. (2017) “Nutritional background changes the hypolipidemic effects of fenofibrate in Nile tilapia (Oreochromis niloticus)”, Scientific Reports, Vol. 7(41706), Nature. https://doi.org/10.1038/srep41706
Norris, D. & Carr, J. A. 2020. Vertebrate Endocrinology, 6th Edition. Academic Press. Cambridge, MA, USA, 656 pp. ISBN: 9780128200933
Pereira, T.S., Boscolo, C.N., Silva, D.G., Batlouni, S.R., Schlenk, D., & Almeida, E.A. (2015). Anti-androgenic activities of diuron and its metabolites in male Nile tilapia (Oreochromis niloticus). Aquatic Toxicology, 164, 10-15. https://doi.org/10.1016/j.aquatox.2015.04.013
Prindiville, J.S. et al. (2011). The fibrate drug gemfibrozil disrupts lipoprotein metabolism in rainbow trout. Toxicology and Applied Pharmacology, Vol. 251, Elsevier, pp. 201-238. doi:10.1016/j.taap.2010.12.013
Runnalls, T.J., Hala, D.N., & Sumpter, J.P. (2007). Preliminary studies into the effects of the human pharmaceutical Clofibric acid on sperm parameters in adult Fathead minnow. Aquatic Toxicology, 84(1), 111-118. https://doi.org/10.1016/j.aquatox.2007.06.005
Sales, C.F., Barbosa Pinheiro, A.P., Ribeiro, Y.M., Weber, A.A., Paes-Leme, F.O., Luz, R.K., Bazzoli, N., Rizzo, E., & Melo, R.M.C. (2020). Effects of starvation and refeeding cycles on spermatogenesis and sex steroids in the Nile tilapia Oreochromis niloticus. Molecular and Cellular Endocrinology, 500, 110643. https://doi.org/10.1016/j.mce.2019.110643
Seki, M., Yokota, H., Matsubara, H., Tsuruda, Y., Maeda, M., Tadokoro, H., & Kobayashi, K. (2002). Effect of ethinylestradiol on the reproduction and induction of vitellogenin and testis-ova in medaka (Oryzias latipes). Environmental toxicology and chemistry, 21(8), 1692–1698.
Selvaraj, S., Ohga, H., Nyuji, M., Kitano, H., Nagano, N., Yamaguchi, A., & Matsuyama, M. (2013). Subcutaneous administration of Kiss1 pentadecapeptide accelerates spermatogenesis in prepubertal male chub mackerel (Scomber japonicus). Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 166(2), 228-36. https://doi.org/10.1016/j.cbpa.2013.06.007
Tang, H., Chen, Y., Wang, L., Yin, Y., Li, G., Guo, Y., Liu, Y., Lin, H., Cheng, C.H.K., & Liu, X. (2018). Fertility impairment with defective spermatogenesis and steroidogenesis in male zebrafish lacking androgen receptor. Biology of Reproduction, 98(2), 227-238. https://doi.org/10.1093/biolre/iox165
Uren-Webster, Tamsyn M, Lewis, Ceri, Filby, Amy L, Paull, Gregory C, & Santos, Eduarda M. (2010). Mechanisms of toxicity of di(2-ethylhexyl) phthalate on the reproductive health of male zebrafish. Aquatic Toxicology, 99(3), 360-369.
Vandermeer, J.H. & Goldberg, D.E. (2013). Population ecology: first principles. Princeton University Press, Princeton, NJ USA.
Velasco-Santamaría, Y.M., Korsgaard, B., Madsen, S.S., & Bjerregaard, P. (2011). Bezafibrate, a lipid-lowering pharmaceutical, as a potential endocrine disruptor in male zebrafish (Danio rerio). Aquatic Toxicology, 105(1-2), 107-118. https://doi.org/10.1016/j.aquatox.2011.05.018
Villeneuve, D. L., Blackwell, B. R., Cavallin, J. E., Collins, J., Hoang, J. X., Hofer, R. N., Houck, K. A., Jensen, K. M., Kahl, M. D., Kutsi, R. N., Opseth, A. S., Santana Rodriguez, K. J., Schaupp, C., Stacy, E. H., & Ankley, G. T. (2023). Verification of In Vivo Estrogenic Activity for Four Per- and Polyfluoroalkyl Substances (PFAS) Identified as Estrogen Receptor Agonists via New Approach Methodologies. Environmental science & technology, 57(9), 3794–3803. https://doi.org/10.1021/acs.est.2c09315
Xia, H., Zhong, C., Wu, X., Chen, J., Tao, B., Xia, X., Shi, M., Zhu, Z., Trudeau, V. L., & Hu, W. (2018). Mettl3 mutation disrupts gamete maturation and reduced fertility in zebrafish. Genetics, 208(2), 729-743. doi: 10.1534/genetics.117.300574
Xie, H., Kang, Y., Wang, S., Zheng, P., Chen, Z., Roy, S., & Zhao, C. (2020). E2f5 is a versatile transcriptional activator required for spermatogenesis and multiciliated cell differentiation in zebrafish. PLoS genetics, 16(3), e1008655. https://doi.org/10.1371/journal.pgen.1008655
Ye, Ting, Kang, Mei, Huang, Qiansheng, Fang, Chao, Chen, Yajie, Shen, Heqing, & Dong, Sijun. (2014). Exposure to DEHP and MEHP from hatching to adulthood causes reproductive dysfunction and endocrine disruption in marine medaka (Oryzias melastigma). Aquatic Toxicology, 146, 115-126.
Zhang, Q., Ye, D., Wang, H., Wang, Y., Hu, W., Sun, Y. (2020). Zebrafish cyp11c1 Knockout Reveals the Roles of 11-ketotestosterone and Cortisol in Sexual Development and Reproduction. Endocrinology, 161(6). https://doi.org/10.1210/endocr/bqaa048