
This AOP is open for adoption and licensed under the BY-SA license. The BY-SA license allows reusers to distribute, remix, adapt, and build upon the material in any medium or format, so long as attribution is given to the creator. The license allows for commercial use. If you remix, adapt, or build upon the material, you must license the modified material under identical terms.
AOP: 459
Title
AhR activation in the thyroid leading to Subsequent Adverse Neurodevelopmental Outcomes in Mammals
Short name
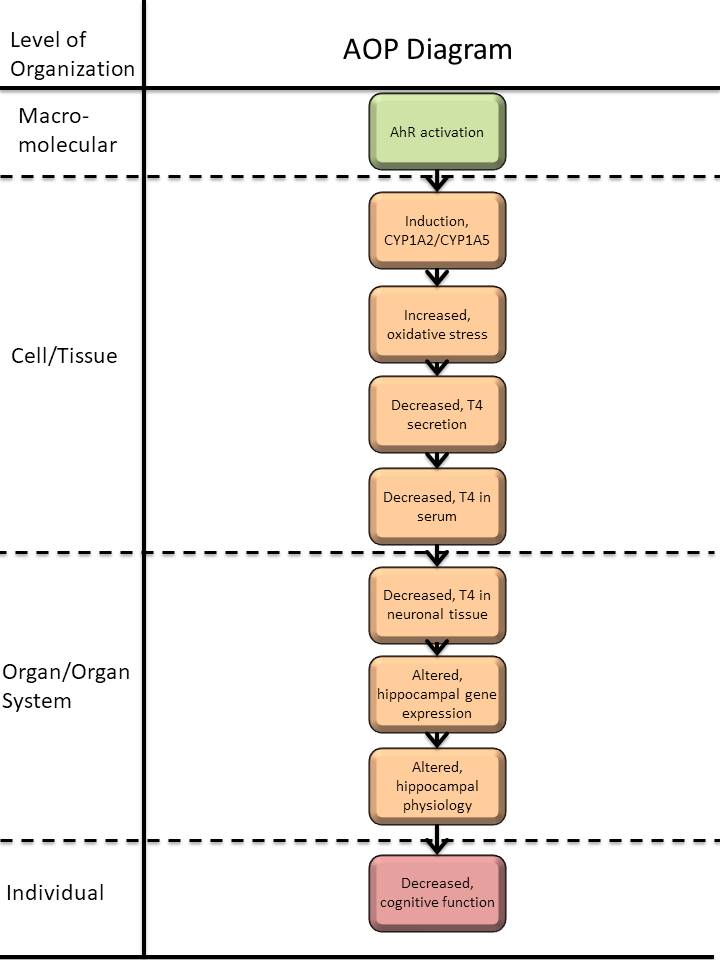
Graphical Representation
Additional AOP Exploration Options
Click links below to explore AOP 459, AhR activation in the thyroid leading to Subsequent Adverse Neurodevelopmental Outcomes in Mammals in tools offered by third parties.
Point of Contact
Contributors
- Prakash Patel
- Simon Thomas
Coaches
OECD Information Table
| OECD Project # | OECD Status | Reviewer's Reports | Journal-format Article | OECD iLibrary Published Version |
|---|---|---|---|---|
This AOP was last modified on July 04, 2024 14:59
Revision dates for related pages
| Page | Revision Date/Time |
|---|---|
| Activation, AhR | May 31, 2025 07:56 |
| Induction, CYP1A2/CYP1A5 | May 30, 2018 14:24 |
| Thyroid hormone synthesis, Decreased | November 04, 2022 09:25 |
| Thyroxine (T4) in serum, Decreased | October 10, 2022 08:52 |
| Thyroxine (T4) in neuronal tissue, Decreased | April 04, 2019 09:13 |
| Hippocampal gene expression, Altered | August 11, 2018 09:26 |
| Hippocampal Physiology, Altered | July 24, 2024 23:15 |
| Cognitive function, decreased | July 25, 2024 17:23 |
| Increase, Oxidative Stress | February 11, 2026 07:05 |
| Activation, AhR leads to Induction, CYP1A2/CYP1A5 | May 07, 2018 08:09 |
| Induction, CYP1A2/CYP1A5 leads to Increase, Oxidative Stress | July 04, 2024 14:32 |
| Increase, Oxidative Stress leads to TH synthesis, Decreased | July 04, 2024 14:13 |
| TH synthesis, Decreased leads to T4 in serum, Decreased | October 10, 2022 08:56 |
| T4 in serum, Decreased leads to T4 in neuronal tissue, Decreased | April 04, 2019 10:50 |
| T4 in neuronal tissue, Decreased leads to Hippocampal gene expression, Altered | August 11, 2018 19:18 |
| Hippocampal gene expression, Altered leads to Hippocampal Physiology, Altered | April 12, 2023 11:28 |
| Hippocampal Physiology, Altered leads to Cognitive function, decreased | July 26, 2024 12:57 |
| 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) | February 09, 2017 14:32 |
| PCB 126 | April 20, 2023 20:20 |
| Polychlorinated biphenyl | November 29, 2016 18:42 |
Abstract
Polychlorinated biphenyls (PCBs) and dioxins are environmental contaminatants whose prenatal exposure in humans and exposure in breast milk is correlated to a specific set of mental impairments (Boucher et al. 2009, Boersma and Lanting 2000, Koopman-Esseboom et al. 1996). What the main mechanism(s) of causing these delays for the various PCBs and dioxins is unclear, but studies in rodents regarding 2,3,7,8 tetrachlorodibenzo-p-dioxin (TCDD) and "dioxin-like" PCBs like PCB-126 that it could be caused by lowering maternal and subsequently foetal thyroxine (T4) levels in the brain.
This AOP summarises the findings that chemicals that are AhR receptor agonists can reduce T4 levels in the plasma in rodents and possibly humans and lead to brain changes in neonates, via the increasing of reactive oxygen species (ROS) in the thyroid reducing the secretion rate of T4 in the thyroid. The problem is that AhR agonists TCDD and dioxin-like PCBs (PCB-126, PCB-169) have multiple effects on several organs as AhR receptor is present in these organs, and especially in the liver whose effect on the liver has been relatively well-characterised (summarised in AOP 458: AhR activation in the liver leading to Subsequent Adverse Neurodevelopmental Outcomes in Mammals).
This AOP was created in order to collate and summarise information around the reduction of T4 levels in the plasma by endocrine disruptors interfering with thyroid hormone secretion via the activation of CYPs. A short summary:
- Chemical activates AhR
- UGT activity in the thyroid increases
- CYP activity in the thyroid increases, generating more reactive oxygen species (ROS)
- ROS affects pathways that reduce secretion of T4 by the thyroid
- Reduction of plasma T4
- Adverse outcomes
AOP Development Strategy
Context
This AOP was developed as part of the ScreenED project, which has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No 825745
Strategy
This AOP summarises the effect of AhR activation leading to thyroid dysfunction, whilst also summarising evidence of hippocampal and mental effects on rodents of TCDD and various PCBs, although the effects on the AhR receptor on the liver metabolism of T4 is probably more dominant. Relevant publications were found by searching through PubMed for terms such as "AhR" "thyroxine" "TCDD" "T4", "CYP", "ROS", "reactive oxygen species", "oxidative stress".
Summary of the AOP
Events:
Molecular Initiating Events (MIE)
Key Events (KE)
Adverse Outcomes (AO)
| Type | Event ID | Title | Short name |
|---|
| MIE | 18 | Activation, AhR | Activation, AhR |
| KE | 850 | Induction, CYP1A2/CYP1A5 | Induction, CYP1A2/CYP1A5 |
| KE | 1392 | Increase, Oxidative Stress | Increase, Oxidative Stress |
| KE | 277 | Thyroid hormone synthesis, Decreased | TH synthesis, Decreased |
| KE | 281 | Thyroxine (T4) in serum, Decreased | T4 in serum, Decreased |
| KE | 280 | Thyroxine (T4) in neuronal tissue, Decreased | T4 in neuronal tissue, Decreased |
| KE | 756 | Hippocampal gene expression, Altered | Hippocampal gene expression, Altered |
| KE | 758 | Hippocampal Physiology, Altered | Hippocampal Physiology, Altered |
| AO | 402 | Cognitive function, decreased | Cognitive function, decreased |
Relationships Between Two Key Events (Including MIEs and AOs)
| Title | Adjacency | Evidence | Quantitative Understanding |
|---|
| Activation, AhR leads to Induction, CYP1A2/CYP1A5 | adjacent | High | Moderate |
| Induction, CYP1A2/CYP1A5 leads to Increase, Oxidative Stress | adjacent | Moderate | Low |
| Increase, Oxidative Stress leads to TH synthesis, Decreased | adjacent | Moderate | Low |
| TH synthesis, Decreased leads to T4 in serum, Decreased | adjacent | High | High |
| T4 in serum, Decreased leads to T4 in neuronal tissue, Decreased | adjacent | High | Moderate |
| T4 in neuronal tissue, Decreased leads to Hippocampal gene expression, Altered | adjacent | Moderate | Moderate |
| Hippocampal gene expression, Altered leads to Hippocampal Physiology, Altered | adjacent | Moderate | Moderate |
| Hippocampal Physiology, Altered leads to Cognitive function, decreased | adjacent | Moderate | Moderate |
Network View
Prototypical Stressors
Life Stage Applicability
| Life stage | Evidence |
|---|---|
| Foetal | High |
| Birth to < 1 month | High |
Taxonomic Applicability
Sex Applicability
| Sex | Evidence |
|---|---|
| Mixed | Moderate |
Overall Assessment of the AOP
Domain of Applicability
Domain of Applicability
- Chemicals: This AOP applies to a wide range of chemicals structures that activate AhR either in vivo or in vitro, but one needs to understand that not all AhR agonists do not produce toxic effects
- Sex: This AOP applies to males and females. Disruption of thyroid hormone regulation during foetal and early postnatal development, but the subsequent adverse impacts on nervous system development may differ and be more severe in males than females (Seo et al. 1999, Rice 1999).
- Life stages: The relevant life stages for this AOP are fetal and early postnatal ages during critical windows of nervous system development where thyroid hormones guide normal development of the brain. There are clear windows of developmental susceptibility and different brain regions show distinct ontogenetic profiles for TH requirements. Distinct phenotypes have been described in both humans and animal models for different periods of TH insufficiency. The influence of maternal thyroid status prior to onset of fetal thyroid function is an important consideration. This AOP does not apply to adult life states.
- Taxonomic: Based on the majority of the available evidence the taxonomic applicability domains of this AOP is mammals. Most evidence for this AOP has been gathered primarily from laboratory rodents and humans.
Essentiality of the Key Events
|
Key Events |
Direct evidence |
Indirect evidence |
No or contradictory evidence |
|
MIE 18 AhR activation |
TCDD and various PCBs can be confirmed to be agonists of AhR from both the EROD (Petrulis et al. 1999) and CALUX assay (Murk et al. 1996) to estimate exposure and activation of the AhR-ARNT pathways. |
||
|
KE 850 Induction, CYP1A2/CYP1A5 |
TCDD – upregulation of CYP1A1 and CYP1A2 in mouse pups heterozygous for AhR +/- gene but far less upregulation of CYPs for pups homozygous for the AhR -/- gene whose mothers were administered 10 ug/kg TCDD on gestation day 12.5 (Nishimura et al. 2005) A marked induction of CYP1A1 mRNA in pups of Holtzmann rat mothers (a TCDD sensitive strain) administered TCDD on gestational day 15 and perinatal exposure to TCDD was found (Nishimura et al. 2003) TCDD – Generally accepted that TCDD and structural analogues induce CYP1A1 and CYP1A2 in animals and humans (Poland and Knutson 1982, Goldstein and Safe 1989, Silbergeld and Gasiewicz 1989 – refs in Tritscher et al. 1992) SD rats were administered TCDD and CYP1A1 and CYP1A2 were induced in liver microsomes in a dose dependent manner (Tritscher et al. 1992) Male F344/NCr rats given a single dose of TCDD or five days of various PCBs increased RNA expression of CYP1A1 and EROD assay activity in liver microsomes (Dragnev et al. 1995) The Cyp1A1 gene was upregulated in Hepa1c1c7 cells and mouse C57BL/6 mouse liver samples on exposure to TCDD and PCB-126 (Nault et al. 2013) Porcine thyrocytes exposed to TCDD and PCB-126 have increased expression of CYP1A1 (Pocar et al. 2006) |
||
|
KE 1392 Oxidative Stress |
ROS production is tightly linked to the catalytic circle of CYP enzymes, more precisely to a phenomenon called “uncoupling”. Briefly, in the presence of NADPH, CYP monooxygenases reduce molecular oxygen to introduce one oxygen equivalent into the substrate and the second one into water. By analysing the stoichiometry of this reaction, it became evident that most CYP enzymes expend more oxygen than needed to monooxygenate the substrates - in fact the small amounts of H2O2 are frequently formed as a by-product of this catalysis (Vogel et al. 2020) TCDD-induced CYPs are a major source of ROS in hepatocytes, as measured in the H2DCFDA assay and the presence of increased 8-oxo-dG (Knerr et al. 2006) Overexpression of CYP1A1 and CYP1A2 in human lymphoblast derived microsomesstimulated production of ROS such as superoxide radicals (Puntanulo and Cedarbaum 1998) TCDD - intraperitoneal treatment of 5ug/kg TCDD in female C57BL/6J mice for three consecutive days produced increased hepatic glutathione levels 2-fold within one week and last for 8 weeks, with urinary levels of hydroxydeoxyguanosine - a product of DNA base oxidation and repair - remain elevated 20-fold for 8 weeks after treatment. (Shertzer et al. 1998) TCDD - when applied to the hepatoma cell line Hepa1c1c7 results in an elevation in the excretion rate of 8-oxoguanine, a biomarker of oxidative DNA damage. (Park et al. 1996) |
TCDD did not increase ROS in HepG2 cells in the H2DCFDA assay or increase 8-oxo-dG (Knerr et al. 2006) |
|
|
KE 277 Thyroid hormone synthesis, Decreased |
In chicken ex vivo thyroids, there was a reduction of T3 and T4 secretion after 24h exposure to TCDD or PCB 126. TCDD and PCB 126 significantly increased mRNA expression of TPO and TG genes, but only PCB 126 reduced NIS genes (Katarzyńska et al. 2015) Chicken thyroid explants were incubated in medium supplemented with TSH (250 mU/ml), PCB118, PCB153, 4-OH-PCB107, and 3-OH-PCB153 (0.5 × 10-8 M), and TSH together with each PCB and OH-PCB. Except for 4-OH-PCB107, all applied PCBs and OH-PCBs inhibited basal and TSH-stimulated T4 secretion. Moreover, they increased basal and reduced TSH-stimulated T3 secretion. (Kowalik and Sechman 2022) Porcine thyrocytes exposed to TCDD and PCB-126 induced a significant downregulation in the expression of NIS and cathepsin B, enzymes involved in the uptake of iodine and release of T4 from thyroglobulin hence T4 secretion (Pocar et al. 2006) |
Hydrogen peroxide is necessary for thyroperoxidase to work, and is most probably generated by DUOX1 and DUOX2. Patients with mutations on DUOX2 gene experience congenital hypothyroidism (Moreno et al. 2002) |
|
|
KE 281 Thyroxine (T4) in serum, Decreased |
There are no other primary sources of T4 in the body so a lack of secretion of T4 by the thyroid would lead to a lack of T4 in the serum. Both PCB-126 and TCDD reduce serum T4 levels in chronic studies in rats (Kohn et al. 1996, NTP 2006), although the dominant effect is likely to be increase in T4 metabolism in the liver rather than secretion by the thyroid (AOP 458) |
||
|
KE 280 Thyroxine (T4) in neuronal tissue, Decreased |
Several studies have demonstrated that foetal brain TH levels, previously decreased by maternal exposure to TH synthesis inhibitors or thyroidectomy, recovered following maternal supply of T4 (e.g. Calvo et al. 1990). However, there are no studies with direct infusion of T4 or T3 directly into brain. |
The upregulation of deiodinase has been shown to compensate for some loss of neuronal T3 (Escobar-Morreale et al. 1995; Escobar-Morreale et al. 1997). Indirect evidence shows that T4 replacement that brings circulating T4 concentration back to physiological levels normal, leads to recovery of brain TH and prevents downstream effects including alterations in cell morphology, differentiation and function. |
PCB-118: lower-TEQ congener (PCB-118) acted via the TH pathway in the rat foetus while the higher-TEQ congener (PCB-126) did not and acted mainly via reducing maternal thyroid hormone levels (Fritsche et al. 2005, Gauger et al. 2004.) |
|
KE 756 Hippocampal gene expression, Altered |
Hippocampal Gene Expression, Altered: It is well established specific genomic pathways underlie the progression of a number of neurodevelopmental processes in the hippocampus. There is some evidence from ex vivo studies that administration of growth factors will reverse the hippocampal dysplasia seen in Jacob/Nsfm knockout mice (Spilker et al., 2016). Less is known about the impact of hormone replacement on TH-responsive gene expression and the qualitative and quantitative relationships between altered TH-dependent gene expression in this brain region and altered hippocampal cytoarchitectural anatomy. |
||
|
KE 758 Hippocampal Physiology, Altered |
Hippocampal anatomy, altered: It is well accepted that normal hippocampal anatomy is critical for hippocampal physiological function, and that alterations in anatomy lead to altered neuronal activity in the hippocampus (Lee et al., 2015; Grant et al., 1992; Spilker et al., 2016). Hippocampal physiology, altered: It is a well-accepted assertion that hippocampal synaptic integrity and neuronal plasticity are essential for spatial information processing in animals and spatial and episodic memory in humans. However, other brain regions also can influence these complex behaviors. Limited data from studies in BDNF knockout animals demonstrate that deficits in hippocampal synaptic transmission and plasticity, and downstream behaviors can be rescued with recombinant BDNF (Aarse et al., 2016; Andero et al., 2014). The relative size of the IIP-MF in females and males. The IIP-MF was smaller in TCDD exposed female AhR-/- mice with respect to their genotype control group. (Powers et al. 2005) |
Pregnant Sprague-Dawley rats were given a consecutive daily dose of TCDD (200 or 800 ng/day/kg) or an equivalent volume of vehicle by gavage on gestational days 8-14 as the prenatal TCDD exposure model. When the male pups grew to adults, morphology and number of neurons in the hippocampus CA1 region was not affected, although the activity of astrocytes in the same region was significantly reduced (Zhang et al. 2018) |
|
|
AO 402 Cognitive Function, Decreased |
TCDD exposed female AhR-/- mice performed worse on the spatial water maze task than non exposed mice (Powers et al. 2005) Pregnant Sprague-Dawley rats were given a consecutive daily dose of TCDD (200 or 800 ng/day/kg) or an equivalent volume of vehicle by gavage on gestational days 8-14 (GD 8-14). The results of the behavioural tests showed that gestational TCDD exposure induced premature motor activity and earlier eyes-opening, but lead to serious deficits of spatial memory and learning ability in the adult male offspring. (Zhang et al 2018) Monkeys exposed to TCDD perinatally exhibited retarded learning of the shape reversals (Shantz and Bowman 1994) |
Pregnant Long-Evans rats were dosed by gavage with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) or at a dose of 0, 200, or 800 ng/kg on gestational day 15, and the offspring was tested during adulthood. Paired-associate learning was found to be impaired in the TCDD-200 and group, but not in either group exposed to 800 ng/kg TCDD (Kakeyama et al. 2014) |
Long-Evans dams dosed at h 0, 0.25, or 1.0 ug/kg/day PCB-126 Monday to Friday beginning 5 weeks before and continuing through gestation and lactation. On the spatial delayed alternation task, there was no convincing evidence for impairment as a result of PCB exposure on adult male pups, as assessed by overall accuracy of performance and measures of perseverative and other types of inappropriate responding (Rice 1999). Time-mated Sprague–Dawley rats were gavaged with eitherTCDD (0.1 mg/kg/day) or corn oil vehicle on gestation days 10–16. On day 80, both male and female TCDD-exposed grown-up pups showed a deficit in learning on the visual partial discrimination-reversal learning task, but TCDD-exposed male rats displayed a pronounced decrease in errors relative to control males in the Morris Water Maze task. There was no difference in the performance of TCDD exposed rats in the radial arm maze task. (Seo et al. 1999) Pregnant Sprague–Dawley rats (10 per dose) received either 0 or 0.1 mg/kg TCDD orally in corn oil from GD 10 to GD 16. One male and one female from each litter were tested beginning at 100 days of age. the results of the current study underscore the fact that (1) alterations in cognitive function observed following early TCDD exposure are very subtle and (2) under some conditions, learning is actually facilitated, rather than impaired, in TCDD-exposed animals. (Widholm et al. 2003) Monkeys exposed to TCDD perinatally showed a slight facilitation of learning on both delayed spatial alternation and spatial reversal learning tasks (Seegal and Schantz 1994) |
Evidence Assessment
Biological plausibility
Biological plausibility refers to the structural or functional relationship between the key events based on our fundamental understanding of "normal biology".
In general, the biological plausibility and coherence linking AhR activation by PCBs and dioxins to decreases in circulating concentrations of THs is great; the biological plausibility of decreases in circulating concentrations of THs, to adverse impacts in the developing hippocampus and subsequent cognitive behaviours is solid (AOP 42), but the evidence via the reduction of thyroid hormone secretion by the thyroid is weak.
Concordance of dose-response relationships:
There are several studies that include correlative evidence between exposure to AhR agonists TCDD (Kohn et al. 1996, NTP 2006) and PCB-126 (NTP, 2006) to downstream KEs up until the reduction of T4 in the plasma (KE 281). Referring to the AOPs 42 and 54, there is ample evidence the reduction of plasma T4 correlates to neurological deficits. Again, the evidence that it’s from activation of AhR leading to the reduction of thyroid hormone secretion by the thyroid is weak.
Consistency:
Uncertainties, inconsistencies, and data gaps:
The effect of thyroid hormone secretion by the thyroid is not routinely measured in animals and humans (Tal et al 1999, Karazynska et al 2016) and especially in the case of the effect of thyroid disruptors. One could gauge this in vivo by measuring the balance of T4 in the plasma and other tissues.
Known Modulating Factors
| Modulating Factor (MF) | Influence or Outcome | KER(s) involved |
|---|---|---|
Quantitative Understanding
Currently, it is poor as T4 secretion of the thyroid in vivo is not routinely measured or calculated in regards to the impact by thyroid disruptors.
Considerations for Potential Applications of the AOP (optional)
References
Aarse J, Herlitze S, Manahan-Vaughan D. The requirement of BDNF for hippocampal synaptic plasticity is experience-dependent. Hippocampus. 2016;26(6):739-5
Andero R, Choi DC, Ressler KJ. (2014). BDNF-TrkB receptor regulation of distributed adult neural plasticity, memory formation, and psychiatric disorders. Prog Mol Biol Transl Sci.122:169-92
Calvo R, Obregón MJ, Ruiz de Oña C, Escobar del Rey F, Morreale de Escobar G. Congenital hypothyroidism, as studied in rats. Crucial role of maternal thyroxine but not of 3,5,3'-triiodothyronine in the protection of the fetal brain. J Clin Invest. 1990;86(3):889-99
Dragnev KH, Nims RW, Fox SD, Lindahl R, Lubet RA. Relative potencies of induction of hepatic drug-metabolizing enzyme genes by individual PCB congeners. Toxicol Appl Pharmacol. 1995;132(2):334-42
Escobar-Morreale HF, Obregón MJ, Escobar del Rey F, Morreale de Escobar G.Replacement therapy for hypothyroidism with thyroxine alone does not ensure euthyroidism in all tissues, as studied in thyroidectomized rats. J Clin Invest. 1995;96(6):2828-38.
Escobar-Morreale HF, Obregón MJ, Hernandez A, Escobar del Rey F, Morreale de Escobar G. (1997). Regulation of iodothyronine deiodinase activity as studied in thyroidectomized rats infused with thyroxine or triiodothyronine. Endocrinology. 138(6):2559-68
Fritsche E, Cline JE, Nguyen NH, Scanlan TS, Abel J. Polychlorinated biphenyls disturb differentiation of normal human neural progenitor cells: clue for involvement of thyroid hormone receptors. Environ Health Perspect. 2005;113(7):871-6
Gauger KJ, Kato Y, Haraguchi K, Lehmler HJ, Robertson LW, Bansal R, Zoeller RT. Polychlorinated biphenyls (PCBs) exert thyroid hormone-like effects in the fetal rat brain but do not bind to thyroid hormone receptors. Environ Health Perspect. 2004;112(5):516-23
Goldstein, J. A., and Safe. S. Mechanism of action and structure-activity relationship for the chlorinated dibenzo-p-dioxins and related compounds. In: R. D. Kimbrough and A. A. Jensen (eds.). Halogenated biphenyls, terphenyls, naphthalenes, dibenzodioxins and related products, pp. 239-293. Amsterdam: Elsevier Science Publishers B. V. (Biomedical Division). 1989
Grant SG, O'Dell TJ, Karl KA, Stein PL, Soriano P, Kandel ER. Impaired long-term potentiation, spatial learning, and hippocampal development in fyn mutant mice. Science. 1992;258(5090):1903-10
Kakeyama M, Endo T, Zhang Y, Miyazaki W, Tohyama C. Disruption of paired-associate learning in rat offspring perinatally exposed to dioxins. Arch Toxicol. 2014;88(3):789-98
Katarzyńska D, Hrabia A, Kowalik K, Sechman A. Comparison of the in vitro effects of TCDD, PCB 126 and PCB 153 on thyroid-restricted gene expression and thyroid hormone secretion by the chicken thyroid gland. Environ Toxicol Pharmacol. 2015;39(2):496-503
Knerr S, Schaefer J, Both S, Mally A, Dekant W, Schrenk D. 2,3,7,8-Tetrachlorodibenzo-p-dioxin induced cytochrome P450s alter the formation of reactive oxygen species in liver cells. Mol Nutr Food Res. 2006;50(4-5):378-84
Kohn MC, Sewall CH, Lucier GW, Portier CJ. A mechanistic model of effects of dioxin on thyroid hormones in the rat. Toxicol Appl Pharmacol. 1996;136(1):29-48
Kowalik K, Sechman A. In vitro effects of polychlorinated biphenyls and their hydroxylated metabolites on the synthesis and metabolism of iodothyronines in the chicken (Gallus domesticus) thyroid gland. Gen Comp Endocrinol. 2022;318:113989
Lee KH, Lee H, Yang CH, Ko JS, Park CH, Woo RS, Kim JY, Sun W, Kim JH, Ho WK, Lee SH. Bidirectional Signaling of Neuregulin-2 Mediates Formation of GABAergicSynapses and Maturation of Glutamatergic Synapses in Newborn Granule Cells of Postnatal Hippocampus. J Neurosci. 2015;35(50):16479-93
Moreno JC, Bikker H, Kempers MJ, van Trotsenburg AS, Baas F, de Vijlder JJ, et al. Inactivating mutations in the gene for thyroid oxidase 2 (THOX2) and congenital hypothyroidism. N Engl J Med. 2002;347(2):95–102
Murk AJ, Legler J, Denison MS, Giesy JP, van de Guchte C, Brouwer A. Chemical-activated luciferase gene expression (CALUX): a novel in vitro bioassay for Ah receptor active compounds in sediments and pore water. Fundam Appl Toxicol. 1996;33(1):149-60
National Toxicology Program. NTP technical report on the toxicology and carcinogenesis studies of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) (CAS No. 1746-01-6) in female Harlan Sprague-Dawley rats (Gavage Studies). Natl Toxicol Program Tech Rep Ser. 2006;(521):4-232
National Toxicology Program. NTP toxicology and carcinogenesis studies of 3,3',4,4',5-pentachlorobiphenyl (PCB 126) (CAS No. 57465-28-8) in female Harlan Sprague-Dawley rats (Gavage Studies). Natl Toxicol Program Tech Rep Ser. 2006;(520):4-246
Nault R, Forgacs AL, Dere E, Zacharewski TR. Comparisons of differential gene expression elicited by TCDD, PCB126, βNF, or ICZ in mouse hepatoma Hepa1c1c7 cells and C57BL/6 mouse liver. Toxicol Lett. 2013;223(1):52-9
Nishimura N, Yonemoto J, Miyabara Y, Fujii-Kuriyama Y, Tohyama C. Altered thyroxin and retinoid metabolic response to 2,3,7,8-tetrachlorodibenzo-p-dioxin in aryl hydrocarbon receptor-null mice. Arch Toxicol. 2005;79(5):260-
Nishimura N, Yonemoto J, Miyabara Y, Sato M, Tohyama C. Rat thyroid hyperplasia induced by gestational and lactational exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Endocrinology. 2003;144(5):2075-83
Park JY, Shigenaga MK, Ames BN. Induction of cytochrome P4501A1 by 2,3,7,8-tetrachlorodibenzo-p-dioxin or indolo(3,2-b)carbazole is associated with oxidative DNA damage. Proc Natl Acad Sci U S A. 1996;93(6):2322-7
Petrulis JR, Bunce NJ. Competitive inhibition by inducer as a confounding factor in the use of the ethoxyresorufin-O-deethylase (EROD) assay. Toxicol Lett. 1999;105(3):251-60
Pocar P, Klonisch T, Brandsch C, Eder K, Fröhlich C, Hoang-Vu C, Hombach-Klonisch S. AhR-agonist-induced transcriptional changes of genes involved in thyroid function in primary porcine thyrocytes. Toxicol Sci. 2006;89(2):408-14
Poland A, Knutson JC. 2,3,7,8-tetrachlorodibenzo-p-dioxin and related halogenated aromatic hydrocarbons: examination of the mechanism of toxicity. Annu Rev Pharmacol Toxicol. 1982;22:517-54
Powers BE, Lin TM, Vanka A, Peterson RE, Juraska JM, Schantz SL. Tetrachlorodibenzo-p-dioxin exposure alters radial arm maze performance and hippocampal morphology in female AhR mice. Genes Brain Behav. 2005;4(1):51-9
Puntarulo S, Cederbaum AI. Production of reactive oxygen species by microsomes enriched in specific human cytochrome P450 enzymes. Free Radic Biol Med. 1998;24(7-8):1324-30
Rice DC. Effect of exposure to 3,3',4,4',5-pentachlorobiphenyl (PCB 126) throughout gestation and lactation on development and spatial delayed alternation performance in rats. Neurotoxicol Teratol. 1999;21(1):59-69
Schantz SL, Bowman RE. Learning in monkeys exposed perinatally to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Neurotoxicol Teratol. 1989;11(1):13-9
Seegal, R. F., and Schantz, S. L. (1994). Neurochemical and behavioral sequelae of exposure to dioxins and PCBs. In Dioxins and Health (A. Schecter, Ed.), pp. 409–447. Plenum Press, New York
Seo BW, Sparks AJ, Medora K, Amin S, Schantz SL. Learning and memory in rats gestationally and lactationally exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Neurotoxicol Teratol. 199; 21(3):231-9
Shertzer HG, Nebert DW, Puga A, Ary M, Sonntag D, Dixon K, Robinson LJ, Cianciolo E, Dalton TP. Dioxin causes a sustained oxidative stress response in the mouse. Biochem Biophys Res Commun. 1998;253(1):44-8
Silbergeld EK, Gasiewicz TA. Dioxins and the Ah receptor. Am J Ind Med. 1989;16(4):455-74
Spilker C, Nullmeier S, Grochowska KM, Schumacher A, Butnaru I, Macharadze T, Gomes GM, Yuanxiang P, Bayraktar G, Rodenstein C, Geiseler C, Kolodziej A, Lopez-Rojas J, Montag D, Angenstein F, Bär J, D'Hanis W, Roskoden T, MikhaylovaM, Budinger E, Ohl FW, Stork O, Zenclussen AC, Karpova A, Schwegler H, Kreutz MR.A Jacob/Nsmf Gene Knockout Results in Hippocampal Dysplasia and Impared BDNF Signaling in Dendritogenesis. PLoS Genet. 2016;12(3):e1005907
Tritscher AM, Goldstein JA, Portier CJ, McCoy Z, Clark GC, Lucier GW. Dose-response relationships for chronic exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin in a rat tumor promotion model: quantification and immunolocalization of CYP1A1 and CYP1A2 in the liver. Cancer Res. 1992;52(12):3436-42
Vogel CFA, Van Winkle LS, Esser C, Haarmann-Stemmann T. The aryl hydrocarbon receptor as a target of environmental stressors - Implications for pollution mediated stress and inflammatory responses. Redox Biol. 2020;34:101530
Widholm JJ, Seo BW, Strupp BJ, Seegal RF, Schantz SL. Effects of perinatal exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin on spatial and visual reversal learning in rats. Neurotoxicol Teratol. 2003;25(4):459-71
Zhang HJ, Liu YN, Xian P, Ma J, Sun YW, Chen JS, Chen X, Tang NJ. Maternal exposure to TCDD during gestation advanced sensory-motor development, but induced impairments of spatial learning and memory in adult male rat offspring. Chemosphere. 2018; 212:678-686